Биофизика - раздел науки, посвященный изучению физ. и физ--хим. явлений
в биол. объектах; её задача - исследование фундам.
процессов, лежащих в основе живой природы. Как самостоят. отрасль науки Б. оформилась
в 1961 (1-й междунар. биофиз. конгресс). Для изучения отд. биол. явлений физ.
идеи и методы использовались значительно раньше. Многие физики начиная с эпохи
Возрождения ставили и решали биол. проблемы, нек-рые физ. задачи были решены
в результате попыток исследовать биол. явления.
Применение физ. идей и
методов к биол. объектам требует учёта их специфики, что и определяет Б. как
самостоят. отрасль науки. Специфика биол. объектов заключается в том, что в
их построении участвует информация, возникшая в результате эволюции и содержащаяся
в наборе генов (геноме). Эта информация проявляется в структуре биол. объектов,
к-рая упорядочена, апериодична, термодинамически неравновесна и приспособлена
для выполнения определ. функции. По структуре биол. объекты аналогичны искусств.
конструкциям (к-рые также строятся целесообразно на основании информации, накопленной
человечеством). Это свойство биол. структур имеет место на всех уровнях: макромолекулярном
(белки, ферменты; см. Полимеры биологические ),клеточном (органеллы и
мембраны; см. Клеточные структуры)и организменном. Существуют два пути
учёта биол. информации, заключённой в объекте: прямой и косвенный.
Первый путь предполагает
построение структуры живого объекта (на всех его уровнях от макромолеку-лярного
до организменного) на основе информации, заложенной в его геноме. В природе
этот путь реализуется вонтогенезе , т. е. процессе развития организма из оплодотворённой
яйцеклетки. При его тео-ретич. исследовании в Б. используют методы теории самоорганизации
(см. Синергетика)и матем. моделирования (см. ниже).
Др. путь - эксперим. исследование
структуры биол. объекта, при этом используют все известные физ. методы. Богатую
информацию на макромолекулярном уровне даёт рентгеновский структурный анализ, на уровне мембран и клеточных органелл - электронная микроскопия ,на
более высоких уровнях - микроскопия и анатомия. Получаемая информация эквивалентна
биол. информации, заложенной в объекте, иными словами, если известна сама конструкция,
то нет необходимости знать информацию, на основе к-рой она была построена. Дальнейшее
исследование поведения объекта проводится в Б. на основании законов физики и
химии с учётом конструкции объекта. В Б. развиваются оба пути, но при решении
конкретных задач второй преобладает.
Согласно принятой классификации, биофизика разделяется на молекулярную
биофизику, клеточную биофизику и биофизику сложных систем. Иногда выделяют в
качестве самостоят. разделов биомеханику, биоэнергетику, матем. биофизику.
Молекулярная биофизика.
В её задачу входит исследование физ. и физ--хим. свойств и взаимодействий макромолекул
и молекулярных комплексов, составляющих живые организмы. Сюда же относятся задачи
определения структуры биол. систем на молекулярном уровне, тесно связанные с
биохимией, а также процессы превращения и миграции энергии. Наиб. важны для
биофизических исследования молекул белков и нуклеиновых K-T.
Белковые макромолекулы представляют собой линейные полимеры, состоящие из цепочки аминокислот. Полимер
свёрнут в структуру (глобулярную либо фибриллярную). Биол. катализаторы (наз.
ферментами или энзимами) имеют глобулярную форму. Последовательность аминокислот
в каждом белке (первичная структура) задаётся генетически; укладка этой цепочки
в глобуле (наз. третичной структурой) определяется первичной структурой и одинакова
во всех молекулах данного белка. Третичную структуру стабилизируют водородные
связи, ван-дер-ваальсовы силы, гидрофобные взаимодействия, а также солевые и
дисульфидные мостики. Выделяют след. элементы
белковой конструкции (наз. вторичными структурами): -спиральные
участки, -структуры
и "шарнирные" группы. Физ. свойства элементов существенно различны.
Так, -спирали представляют
собой жёсткие стержни, в -структурах
первичная последовательность уложена в виде складок. "Шарнирные"
участки содержат малые аминокислоты и допускают повороты жёстких участков. Кроме
того, в белках имеются неспира-лизованные участки, характеризующиеся меньшей
жёсткостью. Нек-рые белки-ферменты состоят из неск. макромолекул, составляющих
т. н. четвертичную структуру.
Непосредств. участие в
биохим. реакциях принимает небольшое число хим. групп фермента, расположенных
в т. н. активном центре. Процесс состоит из след. этапов: 1) сорбция исходных
хим. соединений на активном центре; 2) реакция внутри образовавшегося комплекса;
3) десорбция конечных соединений (продуктов) с тела фермента. Процесс регулируется
веществами, наз. медиаторами или эффекторами. Среди них имеются ингибиторы (тормозящие
реакцию) и активаторы (ускоряющие её). Активаторы, принимающие непосредств.
участие в процессах в активном центре, наз. кофакторами (или коферментами);
возможна также активация путём воздействия на удалённые от активного центра
участки фермента. Ингибиторы делятся на конкурентные (или изостерические), к-рые
связываются с активным центром, и неконкурентные (аллостерические), воздействующие
на тело фермента. Аллостерич. активация и торможение связаны с изменением конструкции
фермента при взаимодействии его с эффекторами.
Конформационные переходы.
Конформацией белка-фермента наз. состояние, в к-ром определена вся конструкция
макромолекулы. Молекулы белка могут находиться в неск. конформациях, в к-рых
топология укладки первичной последовательности и размеры -спиралей
и -структур одинаковы,
но связи между ними различны (а следовательно, различны и конструкции). Переходы
между конформациями, т. н. конформац. переходы (КП), происходят при изменениях
внеш. условий (температуры, влажности и т. п.), зарядового состояния, взаимодействия
с субстратом, медиатором и т. п. Изменения характеристик (ср. размеров, плотности
и т. п.) при КП невелики, но каталитич. способности меняются очень сильно.
Теория КП основана на определении
свободных энергий разных конформаций и аналогична теории фазовых переходов в
физике конденсир. сред. Отличия от этой теории таковы: 1) размеры макромолекулы
ограничены, поэтому переход нерезок: существует область параметров, в к-рой
присутствуют молекулы обеих конформаций; 2) при подсчёте свободной энергии необходимо
учитывать вклад упругой энергии; 3) энтальпийный и энтропийный вклады в свободную
энергию могут быть локализованы в разных частях макромолекулы; 4) относит. изменения
энтальпии и энтропии (отнесённые к массе макромолекулы) при КП могут быть невелики;
КП может происходить в небольшой части конструкции, что тем не менее ведёт к
существ. изменению её характера.
В процессе ферментативного
катализа происходит ряд КП. В биофизике развиты спец. методы, позволяющие определить
число стадий ферментативной реакции и кинетич. коэф. перехода между ними. Скорость
всей ферментативной реакции определяется кинетич. коэф. наиболее медленной стадии.
Осн. проблемой ферментативной кинетики является природа механизмов, обеспечивающих
высокую эффективность и специфичность ферментов. Эффективность означает, что
скорости ферментативных реакций в 107-1010 раз выше скоростей
аналогичных реакций без фермента (т. н. конгруэнтных реакций, проходящих через
те же промежуточные состояния, что и ферментативные). Специфичность фермента
означает, что скорость катализируемой реакции меняется в 103-104
раза при небольшой хим. модификации субстрата. Вопрос о зависимости скорости
ферментативной реакции от концентраций субстратов, ингибиторов и активаторов
решают с помощью матем. моделирования.
Энергетика ферментативного
катализа. В экзоэргич. реакциях источником энергии может служить процесс образования
комплекса за счёт взаимодействия определ. атомов (или групп) субстрата с контактными
группами фермента. Вводят след. энергетич. характеристики: полная энергия Еполн,
равная сумме выделяющихся при образовании контактов энергий, если они взаимодействуют
независимо друг от друга; свободная энергия связывания
; теплота связывания ,
к-рая представляет собой энтальпийную часть .
Разность - энергия
напряжения; величина
- изменение энтропии, где T - абс. темп-pa. Известно неск. схем (моделей)
ферментативного катализа. В модели "ключ-замок" предполагают полную
комплементарность субстрата и фермента. При этом Ен=0 и катализ
имеет энтропийный характер: субстрат в комплексе принимает одну из возможных
конфигураций, благоприятную для послед. реакции.
Аллостерич. эффекты этой
моделью не описываются. В модели "рука - перчатка" считают фермент
достаточно эластичным, что обеспечивает полную комплементарность. При этом EН=0
и катализ имеет энтропийный характер. Модель может описывать аллостерич. эффекты
за счёт изменения эластичности фермента. В модели "дыбы" предполагают,
что фермент абсолютно жёсткий, комплементарность неполная, благодаря чему субстрат
в комплексе напряжён так, что энергия напряжения Ен>0 сосредоточена
на атакуемой связи. Катализ имеет как энтропийный, так и энтальпийный характер.
В модели "белок-машина" в комплексе напряжены как субстрат, так
и фермент. Модель "дыбы" является частным случаем схемы "белок
- машина", когда жёсткость фермента много больше жёсткости субстрата.
В эндоэргич. реакциях (таких,
как синтез АТФ) важен вопрос о том, в какой момент должна быть подана энергия
от стороннего источника. Иногда она необходима лишь для десорбции готового продукта
из комплекса.
Нуклеиновые кислоты, дезоксирибонуклеиновая
(ДНК) и рибонуклеиновая (РНК) к-ты, - линейные полимеры, состоящие из нуклеотидов
четырёх типов, содержащих аденин (А), гуанин (Г), цитозин (Ц) и тимин (T), в
РНК вместо тимина используется урацил (У). Их биол. функции - хранение информации,
передача её потомству, а также реализация информации, записанной в ДНК, при
биосинтезе белков.
В ДНК информация записана
в виде последовательности 4 нуклеотидов, в белках - в форме последовательности
20 аминокислот. Трансляция 4-буквенной записи на 20-буквенную при биосинтезе
белка осуществляется на основе единого для всей биосферы триплетного кода [т.
е. соответствия между триплетами нуклеотидов (к о д о н а м и) и аминокислотами].
Биосинтез белка регулируется
на неск. уровнях: 1) в клеточном ядре синтез информационной РНК (иРНК) на участке
ДНК, несущем информацию об определ. белке, происходит только в том случае, если
нач. участок этого гена (оперон) не заблокирован белками-репрессорами. Последние
синтезируются с участием спец. гена-регулятора, но блокируют оперон лишь в присутствии
корепрессоров - веществ, поступающих в ядро из цитоплазмы. Они передают в ядро
информацию о том, необходим ли синтез данного белка или потребность в нём отпала;
2) уже синтезированные иРНК перед выходом в цитоплазму подвергаются "редактированию",
нек-рые участки из них удаляются спец. белками, а оставшиеся сшиваются.
В результате этих процессов
только малая часть информации, содержащейся в ДНК, одновременно используется
в биосинтезе белка. При изменении условий и (или) в процессе развития организма
происходит переключение биосинтеза; одни участки ДНК блокируются, а другие активируются
(дерепрессируются).
Клеточная биофизика. В
задачу клеточной биофизики входит изучение физ--хим. свойств клетки, функций клеточных
структур, энергетики и термодинамики клеточных процессов, биоэлектрич. процессов.
Структура клетки. Схема
строения клетки изображена на рис. 1, в ней представлены след. клеточные структуры:
клеточная мембрана, отделяющая внутриклеточную среду (цитоплазму) от внешней;
ядро, окружённое ядерной мембраной; митохондрии, отделённые от цитоплазмы спец.
мембраной; комплекс Гольджи, лизосомы, а также более мелкие, не ограниченные
спец. мембраной структуры (рибосомы, микрофиламенты и микротрубочки).
Автономные структуры (ядро,
митохондрии, рибосомы) наз. органеллами; они выполняют след. функции: в ядре
- хранение и транскрипция генетич. информации; в митохондриях - синтез АТФ (см.
ниже); в рибосомах - синтез белка. В фотосинтезирующих клетках растений имеются,
кроме упомянутых органелл, хлоропласты, синтезирующие АТФ за счёт энергии света
(см. Фотосинтез ).В мышечных клетках существуют спец. сократит. структуры.
В низших одноклеточных организмах (прокариотах) ядро отсутствует и генетич.
материал распределён по плазме.
Живая клетка представляет
собой термодинамически неравновесную открытую систему. Это проявляется
в неоднородности пространственного распределения вещества, наличии электрич.
полей и в хим. составе. Концентрации ионов (и др. веществ) в органеллах, в плазме
клетки и во внеш. среде существенно различны, напр. отношение концентраций ионов
H+ может достигать неск. порядков. Различие обеспечивается присутствием
мембран и процессами активного транспорта веществ (т. н. переноса их из области
низкой концентрации в область высокой). Благодаря неравномерному распределению
ионов электрич. потенциалы внеш. среды, цитоплазмы и внутр. среды органелл различны.
Разности потенциалов
10 мВ; градиенты потенциала сосредоточены на соответств. мембранах; поля в них
~104-105 В/см.
Энергетика клетки. В составе
клетки имеются т. н. макроэргич. вещества, чаще всего адено-зинтрифосфорная
к-та (АТФ). При её гидролизе выделяется энергия и АТФ переходит в АДФ (аденозин-дифосфорную
к-ту). Схема реакции:
Величина колеблется
от 0,3 до 0,5 эВ.
Спонтанный гидролиз АТФ
протекает очонь медленно; ферментативный гидролиз, напротив, достаточно быстро;
соответств. ферменты наз. АТФ-азами. Благодаря этим свойствам АТФ выполняет
в клетке роль энергоносителя (или универсальной энергетич. "валюты").
АТФ запасается и хранится в клетке достаточно долго. Гидролизуется АТФ в тех
местах и процессах, где требуется затрата энергии (биосинтез, активный транспорт,
мышечное сокращение и т. п.), т. е. на тех макромолекулах и структурах, к-рые
совершают работу (они же являются АТФ-азами). Энергия гидролиза идет на покрытие
дефицита эндоэргич. реакций.
Синтез АТФ требует затраты
сторонней энергии (равной
), он происходит в след. процессах: неполное окисление глюкозы в цитоплазме;
полное окисление глюкозы (до углекислоты и воды) в митохондриях (окислит. фосфорилирование);
поглощение света в хлоропластах (фотосинтетич. фосфорилирование). Два последних
процесса более эффективны.
Клеточная мембрана. Клетка
может существовать в нескольких функционально разл. состояниях. Переход между
ними регулируется процессами, происходящими, в частности, в клеточной мембране,
к-рая является как бы сенсорным органом клетки, т. е. мембрана воспринимает
сигналы из внеш. среды, преобразует их и передаёт внутр. органеллам.
След. физ. свойства клеточной
мембраны обеспечивают ее регуляторные функции: а) высокая избирательность каналов,
проводящих ионы; существуют разные каналы, каждый проводит преимущественно один
тип ионов (натриевые каналы, калиевые каналы и т. д.); б) каналы могут быть
в активном (проводящем) состоянии и в пассивном. Переход их в активное состояние
(активация) зависит от присутствия в канале ионов - как переносимых, так и сторонних
(другого знака). Сила связывания ионов в канале зависит от электрич. поля (т.
е. от мембранной разности потенциалов ).
Эта зависимость различна для разл. каналов. Поэтому ионный ток через мембрану
является нелинейной функцией величины ,
эта функция может иметь неск. экстремумов; в) в клеточной мембране возможны структурные
переходы (их также наз. конформационными или фазовыми). В них принимают участие
липидный слой мембраны, белковые микротрубочки и микрофиламенты на внутр. поверхности
и полисахаридный слой на внеш. поверхности мембраны. Важную роль играют физ.
свойства системы: механич. целостность внутр. и внеш. оболочек мембраны, их
жёсткость, прочность и т. п. При переходе эти свойства резко меняются, вместе
с ними изменяются вязкость, ионная проводимость и активность мембранных белков-ферментов.
Структурные переходы играют важную роль в управлении делением клеток.
Нервный импульс. Описание
механизма нервного импульса - один из самых ярких примеров использования
физ. идей в биологии. В исходном состоянии внутри клетки имеется избыток ионов
и недостаток
ионов , при этом
внутр. среда заряжена отрицательно по отношению к внешней. Мембранная разность
потенциалов составляет
мВ (для сердечной ткани).
При внеш. воздействии,
ведущем к увеличению
выше порогового значения, -50
мВ, открываются натриевые каналы, возникает пассивный поток ,
что приводит к изменению знака .
По достижении макс. значения 20
мВ натриевые каналы инактивируются полем, поток
компенсируется потоком
и потенциал медленно
уменьшается. В конце этой фазы (характерное время к-рой ~1 мс) калиевые каналы
насыщаются ионами ,
что приводит к резкому увеличению потока
и быстрому падению
до значения -90
мВ. Последний этап (период рефрактерности) - медленное (за время ~1 MC) восстановление
исходного состояния за счёт активации натриевых каналов. Распространение нервного
имлульса связано с электрич. влиянием соседних элементов мембраны друг на друга;
это автоволновой процесс (см. Автоволны).
Клеточный цикл содержит
четыре фазы: G1-период; S-фазу, в к-рой происходит
синтез ДНК и генетич. материал удваивается; -период;
фазу митоза M, в к-рой происходит деление клетки. Схема цикла представлена
на рис. 2. Состояние, в к-ром клетка периодически проходит упомянутые фазы,
наз. пролиферацией. Кроме того, существуют состояния покоя
и, в к-рых клетка
может находиться и функционировать сколь угодно долго. Большинство клеток сложных
организмов находятся в состоянии покоя; для перехода их к пролиферации необходимы
внешние (по отношению к клетке) стимулы.
Механизм стимуляции заключается
в том, что воздействие вызывает структурный (фазовый) переход в клеточной
мембране. Переход может быть вызван как неспецифич. воздействием на внеш. сторону
мембраны (напр., механическим, электрическим и т. д.), так и специфическим (напр.,
гормональным). Гормоны образуют комплексы с соответств. комплементарными им
рецепторами на поверхности клеточной мембраны; вязкоупругие свойства её при
этом изменяются, что вызывает структурный переход. В результате структурного
перехода становится иным состав цитоплазмы, в ней повышается концентрация циклической
аденозинмоно-фосфорной к-ты, что является внутр. сигналом для перехода к пролиферации.
Такой механизм регуляции
обеспечивает стандартизацию отклика (на разл. внеш. воздействия клетка отвечает
одинаково) и возможность варьировать чувствительность к внеш. сигналам в широком
диапазоне. Нарушение управляющего механизма (напр., механич. целостности внеш.
или внутр. оболочек) может привести к тому, что клетка перестанет нуждаться
во внеш. стимулах. Такой неуправляемый режим деления характерен для злокачеств.
клеток. Матем. модель регуляции клеточного цикла (как и модель нервного импульса)
относится к классу релаксац. автоколебат. моделей с N-образной характеристикой
(см. ниже).
Биофизика сложных систем.
В задачу этого раздела входят описание эволюц. процессов, включая возникновение
жизни и развитие организма (см. Эволюция биологическая ),изучение управления
биол. системами (на всех уровнях от молекулярного до экологического) и биомеханика.
Управление биол. системами и их эволюция имеют много общего, они содержат одинаковые
явления: автоколебания ,автоволны, диссипативпые структуры, и
др. Для их описания используют метод матем. моделирования с помощью кинетич.
ур-ний. Имеются два подхода: первый основан на теории марковских случайных
процессов; составляют линейные ур-ния для вероятности Pi застать
систему в определенном i-м состоянии
кинетич. коэф.
предполагаются заданными.
Рис. 2.
Схема клеточного цикла.
Второй подход основан на
теории динамических систем; переменными являются концентрации, числа
особей в экологич. системе, электрич. мембранные потенциалы и т. п. Ур-ния обычно
нелинейны и имеют ту же форму, что и ур-ния хим. кинетики:
Перейти от ур-ний типа
(1) к ур-ниям типа (2) можно, заменяя распределения Pi первыми
моментами. Так, если Pi - вероятность застать в системе объёма
V определ. число ni молекул данного вещества, то его
концентрация равна:
. Ур-ния для моментов, полученные из (1), имеют форму (2), но содержат, помимо
динамич. функции ,
стохастич. добавку. Последняя мала, если распределения достаточно
узки (близки к -образным).
Первый подход используют,
если множество состояний дискретно и число их невелико, напр. в кинетике ферментативных
реакций. При этом
- вероятность застать ферментный комплекс в i-м состоянии. Вероятностный
подход применяют также при описании изменений состояния клетки от момента её
появления до деления (при этом Pi - вероятность застать клетку
в i-й фазе) и в ряде др. задач.
Динамич. подход конструктивен,
когда число состояний системы достаточно велико. Ур-ния записывают на основании
данных о структуре системы, свойствах процесса и характерных временах его стадий.
При этом используют методы редукции системы (т. е. сведения её к системе с меньшим
числом ур-ний и переменных), основанные на принципах временной иерархии и структурной
организации. Временная иерархия означает, что характерные времена процессов
разбиваются на группы так, что внутри группы они одного порядка, но сильно отличаются
от времён других групп. Такая ситуация обычно реализуется в биол. процессах,
поскольку при этом существенно упрощается управление процессом (в т. ч. и самоуправление).
При моделировании процесса с характерным временем временная иерархия позволяет
все переменные с большими временами изменения считать пост. параметрами, а перем.
процессы с меньшими временами выразить через искомые переменные.
Структурная организация
означает, что система разбивается на ячейки, почти (но не полностью) изолированные
друг от друга. Это позволяет поддерживать термодинамически неравновесное состояние
биол. системы, а также упрощает управление ею (и самоуправление). Такая организация
в первом приближении даёт возможность рассматривать процессы в каждой ячейке
независимо и в след. приближении учитывать взаимодействия между ними. Задачи
о движении веществ в пространстве сводятся к обмену между ячейками, к-рый описывается
ур-ниями типа (2).
Математическоемоделирование преследует две цели: 1) качеств. описание нетривиальных явлений, таких,
как автоколебания, возникновение и исчезновение стационарных состояний и т.
п. Для этой цели строят максимально упрощённые (базовые) модели. Большинство
из них состоит из двух ур-ний; 2) количеств. описание конкретных процессов,
качеств. поведение к-рых известно. Для этой цели строят т. н. имитац. модели;
они могут содержать много ур-ний и параметров, к-рые определяют из сравнения
с эксперим. данными.
Модель Лотки и Вольтерра
для описания сосуществования хищников (их число N1) и жертв
(их число N2):
- коэф. рождаемости (принято, что для рождения хищника ему необходимо съесть
жертву), и
- коэф. смертности (принято, что жертвы погибают при встрече с хищником).
Система (3) имеет периодич.
решения и стационарное решение типа центра, она структурно неустойчива (см.
Устойчивость движения)и потому не может описывать реальные процессы.
При небольших модификациях (учёт зависимостей
от N1, N2) она становится структурно устойчивой,
имеет автоколебат. решения и широко используется в экологии.
Модель Гаузе описывает
взаимодействие популяций:
где -
численность i-й популяции, п - число популяций, -
коэф. размножения (разности коэф. рождения и естеств. смертности),
- коэф. взаимодействия, учитывающие конкуренцию за питание, взаимное уничтожение,
эффект тесноты и т. п.
Модель (4) описывает как
отбор "наилучшей" популяции (и исчезновение конкурентов), так и
выбор одной из равноправных. Последнее имеет место, если коэф.
одинаковы, а коэф.
. Выбор оказывается возможным, поскольку симметричное стационарное состояние
при упомянутых условиях неустойчиво. В этом процессе возникает биол. информация.
Модели типа (4) используют в теории биол. эволюции (включая происхождение жизни)
и в экологии. В инженерной микробиологии и теории иммунной реакции организма
используют близкие к (4) модели, но при этом учитывают зависимость коэф.
от времени.
В физиологии популярны
модели, описывающие автоколебат. процессы. В этих процессах реакция объекта
на внеш. воздействие зависит от фазы. Поэтому моделирование важно для определения
оптим. момента воздействия (в т. ч. лекарственного).
В кинетике ферментативных
систем используют модели, в к-рых динамич. переменными являются концентрации
субстратов и продуктов. Нелинейные зависимости скоростей реакций от концентраций
субстратов и медиаторов обеспечивают наличие в системе как положит., так и отрицат.
обратной связи. Модели мембранных процессов строят аналогично, при этом обратные
связи обеспечиваются за счёт нелинейной зависимости скорости транспорта от концентраций.
Благодаря положит. обратной связи (в частности, автокатализу) стационарные состояния
могут терять устойчивость (как и в теории горения), что позволяет описывать
ряд нетривиальных явлений.
Релаксац. модель с N-образной
характеристикой:
где параметр ,
а функция P (х, у)такова, что изоклина y1(X)(решение
ур-ния Р(х, у)=0)имеет два экстремума. Модель описывает генерацию стандартного
сигнала в ответ на малое, но конечное внеш. воздействие и релаксац. автоколебания.
При изменении параметров модели (5) режим "ждущего" стационарного
состояния переходит в режим автоколебаний (и обратно). Модель (5) используют
при описании генерации нервного импульса, возникновения биол. ритмов (т. н.
биол. часов), в теории мембранной регуляции клеточного цикла и моделировании
др. явлений.
Мультистационарная модель
с перем. числом стационарных состояний, напр. система, описывающая переключения
генетич. аппарата с одного режима работы на другой:
В зависимости от параметров
Ax, Ау и
система (6) может иметь либо одно (устойчивое) стационарное состояние, либо
три (два устойчивых и одно неустойчивое). В последнем случае система (6) при
заданном наборе параметров способна функционировать в двух разных режимах.
Переключение из одного состояния в другое возможно при изменении динамич. переменных
х и у за счёт внеш. сил (силовое переключение) и за счёт временного
изменения параметров с последующим возвращением их к исходным значениям (параметрич.
переключение). Модель (6) используют для описания дифференциации клеток при
эволюции организма и для исследования возможностей параметрич. управления онтогенезом.
Нелинейные уравнения матем.
физики (т. н. диффузионно-реакционные) применяют при моделировании возникновения
пространственной структурной организации (самоорганизации), а также возникновения
и распространения импульсов возбуждения.
Самоорганизация (см. Синергетика)в пространстве описывается на основе теории диссипативных структур. Биол.
примерами её являются: а) образование сложного организма из оплодотворённой
яйцеклетки (т. е. процесс морфогенеза). Задача биофизики - выяснить механизмы реализации
генетич. информации о пространственной структуре организма и его органов в процессе
развития организма (онтогенеза). В рамках теории диссипативных структур эта
задача сводится к параметрич. управлению и выяснению условий, при к-рых возникает
единств. структура при заданных (предопределённых генетически) параметрах; б)
образование экологич. структур; предварит. информация о структуре отсутствует,
она сама возникает при образовании вида. Задача биофизики - проследить образование устойчивой
структуры при изменении параметров, граничных и начальных условий.
Возбуждение и распространение
импульсов и волн возбуждения описывается теорией автоволновых процессов. В биологии
к ним относятся: распространение нервных импульсов, перистальтич. волны в кишечнике
и т. п. (при этом используют теорию автоволн в одномерном пространстве); распространение
волн возбуждения в сердечной мышце, в коре головного мозга, сетчатке глаза и
т. п. (при этом применяют теорию автоволн в дву- и трёхмерных пространствах,
к-рая помогает описать и выяснить механизм ряда патологич. явлений).
Для возникновения диссипативных
структур и автоволн необходимо наличие как положит. обратной связи (автокатализ),
так и отрицательной (демпфирование или ингибирование). Эти условия обеспечиваются
за счёт нелинейных зависимостей скоростей ферментативных реакций от субстрата
и скоростей ионного транспорта от электрич. поля.
Биомеханика состоит
из 3 частей: механики макроскопич. движений организма; гидродинамики кровообращения
и внеш. дыхания; механики мышечного сокращения. Биомеханика возникла раньше
др. областей биофизики. Так, изучение механики движения и кровообращения началось задолго
до появления биофизики. как самостоят. науч. направления. [Задача о движении жидкости
по цилиндрич. трубам была поставлена и решена Ж. Л. М.Пуазёйлем (J. L. M, Poiseuille)
в 1840 для описания движения крови по сосудам.]
Специфика биомеханики связана
с важной ролью регуляторных процессов, обеспечивающих обратные связи. Благодаря
этому механич. (или гидродинамич.) параметры (тип конструкции, вязкость жидкости,
размеры сосудов, жёсткость и т. п.), к-рые в механике принимаются постоянными,
в биомеханике могут зависеть от состояния системы.
Так, скелет представляет
собой конструкцию со многими степенями свободы. Система мышц и программа их
упорядоченных во времени сокращений накладывает ограничения, выделяющие одну
степень свободы, именно ту, к-рая наиболее приспособлена для выполнения необходимой
в данный момент функции. Аналогичные искусств. конструкции многоцелевого назначения
уступают реализованным в живой природе. Элементы биол. макроконструкций (т.
е. кости и хрящи скелета животных, стебли растений и т. д.) также обладают спецификой:
эти элементы механически гетерогенны и построены из анизотропных "материалов".
Эта особенность обеспечивает биол. конструкциям высокую прочность при миним.
затратах материала.
Биомеханика периодич. (в
частности, перистальтич.) движений органов связана, в первую очередь, с деятельностью
биол. насосов - сердца, лёгких и тонкого кишечника. К специфике биол. насосов
можно отнести то, что их стенки состоят из мышечной ткани и способны к периодич.
сокращению (что и обеспечивает перекачку). Кроме того, деятельность насосов
регулируется нервными импульсами, поступающими из организма.
Биомеханика кровеносной
и дыхат. систем описывает процессы газообмена (снабжение организма кислородом
и удаление из него углекислоты). Специфика её в следующем: кровь по свойствам
существенно отличается от ньютоновской жидкости, поэтому течение её по сосудам
не описывается ур-нием Пуазёйля; при движении крови по капиллярам (микроциркуляция)
эффективная вязкость и др. параметры не постоянны, а зависят от скорости оксигенации
(дезоксигенации) гемоглобина и др. процессов; при движении дыхат. газов в ветвящейся
бронхиально-альвеолярной системе поверхностное натяжение альвеол не остаётся
постоянным, а регулируется организмом в зависимости от его потребностей.
Биомеханика мышечного сокращения
включает молекулярные процессы сокращения мышечного волокна и управления ими.
Мышечное волокно содержит фибриллярные (нитевидные) белки, к-рые могут скользить
относительно друг друга. Структура их (см. Клеточные структуры,)такова,
что имеется одна выделенная степень свободы, вдоль к-рой и происходит скольжение.
Работа совершается мышцей за счёт гидролиза АТФ. Управление сокращением мышц
осуществляется нервными (или в экспериментах электрическими) импульсами, к-рые
инициируют сокращение. В гладких мышцах сокращение вызывается волной возбуждения
в самой мышечной ткани. Механизм её возникновения и распространения описывается
теорией автоволн. В летательных мышцах насекомых периодич. сокращение происходит
с частотой ~102 Гц и представляет собой автоко-лебат. процесс. При
этом спец. внеш. стимула для каждого сокращения не требуется, управление осуществляется
за счёт воздействия нервных импульсов на параметры автоколебаний.
Осн. нерешёнными задачами биофизики являются проблемы эволюции биосферы (включая возникновение жизни, см. Эволюция
биологическая): кол-во ценной информации, возникающей на разл. этапах эволюции,
и механизм её появления, происхождение генетич. кода и т. п. Эти вопросы важны
и для теории др. развивающихся и самоорганизующихся систем (языки, социальные
структуры и т. п.).
Исходной особенностью применения
физ. идей в биологии является след. принцип: все явления, в т. ч. биологические,
подчиняются осн. физ. законам. В физике накоплен опыт и развиты методы описания
сложных систем, при этом часто используют предположения, упрощающие расчёты
(и применительно к физ. явлениям оправданные). Так, в случае глобальной неустойчивости
механич. систем оправдано предположение о молекулярном хаосе (равносильное эргодической
гипотезе), следствием к-рого является термодинамика равновесных процессов.
В физике твёрдого тела часто прибегают к методам усреднения, основанным на предположении
о микрооднородности объекта. Но попытки использовать в биофизике метод, оправдавший
себя в физике, как правило, не ведут к успеху. Успешным в биофизике является иной путь,
состоящий из двух этапов: 1) анализа реальной структуры биол. объекта (она в
целом неоднородна) и построения на его основе физ. модели, адекватной объекту,
при атом учитывается заключённая в объекте информация и, следовательно, биол.
специфика; 2) анализа модели с использованием известных положений физики
(в т. ч. термодинамики, механики,
гидродинамики) применительно к тем деталям модели, где последние оправданы.
Этот подход характерен для Б на всех ее уровнях: от молекулярного до биосферы
в целом.
Литература по биофизике
Блюмрнфельд Л. А., Пробпсмы биологической филши, 2 изд , M , 1977,
Волькенштейн М. В. Молекулярная биофизика, M.. 1975,
Волькенштейн М. В. Общая биофизика, M , 1978,
Волькенштейн М. В. Биофизика, M, 1981,
Романовский Ю. M., Степанова H. В., Чернявский Д. С. Математическое моделирование в биофизике, M , 1975,
Романовский Ю. M., Степанова H. В., Чернявский Д. С. Математическая биофизика, M, 1984,
Иваницкий Г. P., Кринский В. И., Сельков Е. E., Математическая биофизика клетки, M , 1978
Знаете ли Вы, в чем ложность понятия "физический вакуум"?
Физический вакуум - понятие релятивистской квантовой физики, под ним там понимают низшее (основное) энергетическое состояние квантованного поля, обладающее нулевыми импульсом, моментом импульса и другими квантовыми числами. Физическим вакуумом релятивистские теоретики называют полностью лишённое вещества пространство, заполненное неизмеряемым, а значит, лишь воображаемым полем. Такое состояние по мнению релятивистов не является абсолютной пустотой, но пространством, заполненным некими фантомными (виртуальными) частицами. Релятивистская квантовая теория поля утверждает, что, в согласии с принципом неопределённости Гейзенберга, в физическом вакууме постоянно рождаются и исчезают виртуальные, то есть кажущиеся (кому кажущиеся?), частицы: происходят так называемые нулевые колебания полей. Виртуальные частицы физического вакуума, а следовательно, он сам, по определению не имеют системы отсчета, так как в противном случае нарушался бы принцип относительности Эйнштейна, на котором основывается теория относительности (то есть стала бы возможной абсолютная система измерения с отсчетом от частиц физического вакуума, что в свою очередь однозначно опровергло бы принцип относительности, на котором постороена СТО). Таким образом, физический вакуум и его частицы не есть элементы физического мира, но лишь элементы теории относительности, которые существуют не в реальном мире, но лишь в релятивистских формулах, нарушая при этом принцип причинности (возникают и исчезают беспричинно), принцип объективности (виртуальные частицы можно считать в зависимсоти от желания теоретика либо существующими, либо не существующими), принцип фактической измеримости (не наблюдаемы, не имеют своей ИСО).
Когда тот или иной физик использует понятие "физический вакуум", он либо не понимает абсурдности этого термина, либо лукавит, являясь скрытым или явным приверженцем релятивистской идеологии.
Понять абсурдность этого понятия легче всего обратившись к истокам его возникновения. Рождено оно было Полем Дираком в 1930-х, когда стало ясно, что отрицание эфира в чистом виде, как это делал великий математик, но посредственный физик Анри Пуанкаре, уже нельзя. Слишком много фактов противоречит этому.
Для защиты релятивизма Поль Дирак ввел афизическое и алогичное понятие отрицательной энергии, а затем и существование "моря" двух компенсирующих друг друга энергий в вакууме - положительной и отрицательной, а также "моря" компенсирующих друг друга частиц - виртуальных (то есть кажущихся) электронов и позитронов в вакууме.
Однако такая постановка является внутренне противоречивой (виртуальные частицы ненаблюдаемы и их по произволу можно считать в одном случае отсутствующими, а в другом - присутствующими) и противоречащей релятивизму (то есть отрицанию эфира, так как при наличии таких частиц в вакууме релятивизм уже просто невозможен). Подробнее читайте в FAQ по эфирной физике.
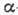 -спиральные
участки,
-спиральные
участки, 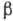 -структуры
и "шарнирные" группы. Физ. свойства элементов существенно различны.
Так,
-структуры
и "шарнирные" группы. Физ. свойства элементов существенно различны.
Так, 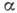 -спирали представляют
собой жёсткие стержни, в
-спирали представляют
собой жёсткие стержни, в 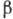 -структурах
первичная последовательность уложена в виде складок. "Шарнирные"
участки содержат малые аминокислоты и допускают повороты жёстких участков. Кроме
того, в белках имеются неспира-лизованные участки, характеризующиеся меньшей
жёсткостью. Нек-рые белки-ферменты состоят из неск. макромолекул, составляющих
т. н. четвертичную структуру.
-структурах
первичная последовательность уложена в виде складок. "Шарнирные"
участки содержат малые аминокислоты и допускают повороты жёстких участков. Кроме
того, в белках имеются неспира-лизованные участки, характеризующиеся меньшей
жёсткостью. Нек-рые белки-ферменты состоят из неск. макромолекул, составляющих
т. н. четвертичную структуру.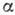 -спиралей
и
-спиралей
и 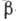 -структур одинаковы,
но связи между ними различны (а следовательно, различны и конструкции). Переходы
между конформациями, т. н. конформац. переходы (КП), происходят при изменениях
внеш. условий (температуры, влажности и т. п.), зарядового состояния, взаимодействия
с субстратом, медиатором и т. п. Изменения характеристик (ср. размеров,
-структур одинаковы,
но связи между ними различны (а следовательно, различны и конструкции). Переходы
между конформациями, т. н. конформац. переходы (КП), происходят при изменениях
внеш. условий (температуры, влажности и т. п.), зарядового состояния, взаимодействия
с субстратом, медиатором и т. п. Изменения характеристик (ср. размеров, 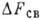 ; теплота связывания
; теплота связывания 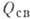 ,
к-рая представляет собой энтальпийную часть
,
к-рая представляет собой энтальпийную часть 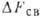 .
Разность
.
Разность 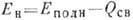 - энергия
напряжения; величина
- энергия
напряжения; величина 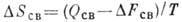 - изменение энтропии, где T - абс. темп-pa. Известно неск. схем (моделей)
ферментативного катализа. В модели "ключ-замок" предполагают полную
комплементарность субстрата и фермента. При этом Ен=0 и катализ
имеет энтропийный характер: субстрат в комплексе принимает одну из возможных
конфигураций, благоприятную для послед. реакции.
- изменение энтропии, где T - абс. темп-pa. Известно неск. схем (моделей)
ферментативного катализа. В модели "ключ-замок" предполагают полную
комплементарность субстрата и фермента. При этом Ен=0 и катализ
имеет энтропийный характер: субстрат в комплексе принимает одну из возможных
конфигураций, благоприятную для послед. реакции. 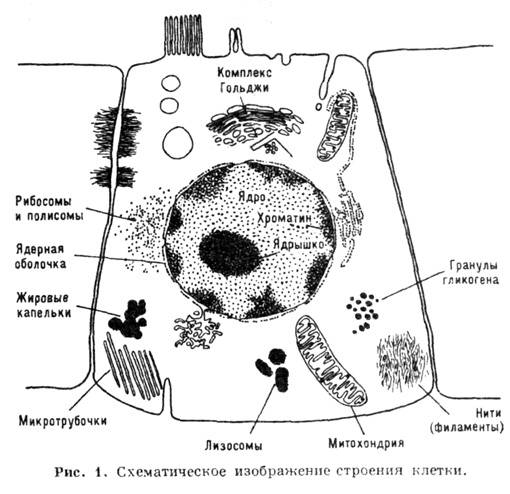
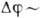 10 мВ; градиенты потенциала сосредоточены на соответств. мембранах; поля в них
~104-105 В/см.
10 мВ; градиенты потенциала сосредоточены на соответств. мембранах; поля в них
~104-105 В/см.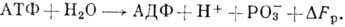
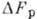 колеблется
от 0,3 до 0,5 эВ.
колеблется
от 0,3 до 0,5 эВ.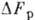 ), он происходит в след. процессах: неполное окисление глюкозы в цитоплазме;
полное окисление глюкозы (до углекислоты и воды) в митохондриях (окислит. фосфорилирование);
), он происходит в след. процессах: неполное окисление глюкозы в цитоплазме;
полное окисление глюкозы (до углекислоты и воды) в митохондриях (окислит. фосфорилирование);
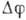 ).
Эта зависимость различна для разл. каналов. Поэтому ионный ток через мембрану
является нелинейной функцией величины
).
Эта зависимость различна для разл. каналов. Поэтому ионный ток через мембрану
является нелинейной функцией величины 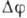 ,
эта функция может иметь неск. экстремумов; в) в клеточной мембране возможны структурные
переходы (их также наз. конформационными или фазовыми). В них принимают участие
липидный слой мембраны, белковые микротрубочки и микрофиламенты на внутр. поверхности
и полисахаридный слой на внеш. поверхности мембраны. Важную роль играют физ.
свойства системы: механич. целостность внутр. и внеш. оболочек мембраны, их
жёсткость, прочность и т. п. При переходе эти свойства резко меняются, вместе
с ними изменяются вязкость, ионная проводимость и активность мембранных белков-ферментов.
Структурные переходы играют важную роль в управлении делением клеток.
,
эта функция может иметь неск. экстремумов; в) в клеточной мембране возможны структурные
переходы (их также наз. конформационными или фазовыми). В них принимают участие
липидный слой мембраны, белковые микротрубочки и микрофиламенты на внутр. поверхности
и полисахаридный слой на внеш. поверхности мембраны. Важную роль играют физ.
свойства системы: механич. целостность внутр. и внеш. оболочек мембраны, их
жёсткость, прочность и т. п. При переходе эти свойства резко меняются, вместе
с ними изменяются вязкость, ионная проводимость и активность мембранных белков-ферментов.
Структурные переходы играют важную роль в управлении делением клеток.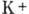 и недостаток
ионов
и недостаток
ионов 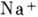 , при этом
внутр. среда заряжена отрицательно по отношению к внешней. Мембранная разность
потенциалов составляет
, при этом
внутр. среда заряжена отрицательно по отношению к внешней. Мембранная разность
потенциалов составляет 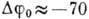 мВ (для сердечной ткани).
мВ (для сердечной ткани).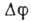 выше порогового значения,
выше порогового значения, 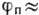 -50
мВ, открываются натриевые каналы, возникает пассивный поток
-50
мВ, открываются натриевые каналы, возникает пассивный поток 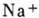 ,
что приводит к изменению знака
,
что приводит к изменению знака 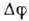 .
По достижении макс. значения
.
По достижении макс. значения 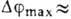 20
мВ натриевые каналы инактивируются полем, поток
20
мВ натриевые каналы инактивируются полем, поток 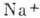 компенсируется потоком
компенсируется потоком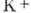 и потенциал
и потенциал 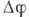 медленно
уменьшается. В конце этой фазы (характерное время к-рой ~1 мс) калиевые каналы
насыщаются ионами
медленно
уменьшается. В конце этой фазы (характерное время к-рой ~1 мс) калиевые каналы
насыщаются ионами 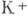 ,
что приводит к резкому увеличению потока
,
что приводит к резкому увеличению потока 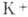 и быстрому падению
и быстрому падению 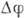 до значения
до значения 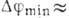 -90
мВ. Последний этап (период рефрактерности) - медленное (за время ~1 MC) восстановление
исходного состояния за счёт активации натриевых каналов. Распространение нервного
имлульса связано с электрич. влиянием соседних элементов мембраны друг на друга;
это автоволновой процесс (см.
-90
мВ. Последний этап (период рефрактерности) - медленное (за время ~1 MC) восстановление
исходного состояния за счёт активации натриевых каналов. Распространение нервного
имлульса связано с электрич. влиянием соседних элементов мембраны друг на друга;
это автоволновой процесс (см. 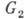 -период;
фазу митоза M, в к-рой происходит деление клетки. Схема цикла представлена
на рис. 2. Состояние, в к-ром клетка периодически проходит упомянутые фазы,
наз. пролиферацией. Кроме того, существуют состояния покоя
-период;
фазу митоза M, в к-рой происходит деление клетки. Схема цикла представлена
на рис. 2. Состояние, в к-ром клетка периодически проходит упомянутые фазы,
наз. пролиферацией. Кроме того, существуют состояния покоя 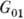 и
и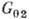 , в к-рых клетка
может находиться и функционировать сколь угодно долго. Большинство клеток сложных
организмов находятся в состоянии покоя; для перехода их к пролиферации необходимы
внешние (по отношению к клетке) стимулы.
, в к-рых клетка
может находиться и функционировать сколь угодно долго. Большинство клеток сложных
организмов находятся в состоянии покоя; для перехода их к пролиферации необходимы
внешние (по отношению к клетке) стимулы.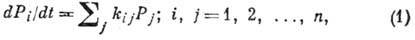
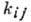 предполагаются заданными.
предполагаются заданными.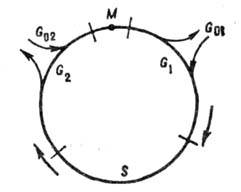

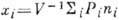 . Ур-ния для моментов, полученные из (1), имеют форму (2), но содержат, помимо
динамич. функции
. Ур-ния для моментов, полученные из (1), имеют форму (2), но содержат, помимо
динамич. функции 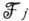 ,
стохастич. добавку. Последняя мала, если распределения
,
стохастич. добавку. Последняя мала, если распределения 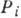 достаточно
узки (близки к
достаточно
узки (близки к 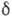 -образным).
-образным).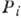 - вероятность застать ферментный комплекс в i-м состоянии. Вероятностный
подход применяют также при описании изменений состояния клетки от момента её
появления до деления (при этом Pi - вероятность застать клетку
в i-й фазе) и в ряде др. задач.
- вероятность застать ферментный комплекс в i-м состоянии. Вероятностный
подход применяют также при описании изменений состояния клетки от момента её
появления до деления (при этом Pi - вероятность застать клетку
в i-й фазе) и в ряде др. задач.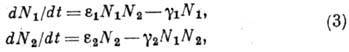
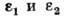 - коэф. рождаемости (принято, что для рождения хищника ему необходимо съесть
жертву),
- коэф. рождаемости (принято, что для рождения хищника ему необходимо съесть
жертву), 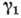 и
и 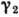 - коэф. смертности (принято, что жертвы погибают при встрече с хищником).
- коэф. смертности (принято, что жертвы погибают при встрече с хищником).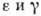 от N1, N2) она становится структурно устойчивой,
имеет автоколебат. решения и широко используется в экологии.
от N1, N2) она становится структурно устойчивой,
имеет автоколебат. решения и широко используется в экологии.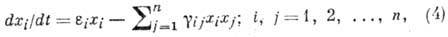
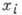 -
численность i-й популяции, п - число популяций,
-
численность i-й популяции, п - число популяций, 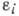 -
коэф. размножения (разности коэф. рождения и естеств. смертности),
-
коэф. размножения (разности коэф. рождения и естеств. смертности), 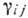 - коэф. взаимодействия, учитывающие конкуренцию за питание, взаимное уничтожение,
эффект тесноты и т. п.
- коэф. взаимодействия, учитывающие конкуренцию за питание, взаимное уничтожение,
эффект тесноты и т. п.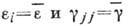 одинаковы, а коэф.
одинаковы, а коэф. 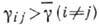 . Выбор оказывается возможным, поскольку симметричное стационарное состояние
при упомянутых условиях неустойчиво. В этом процессе возникает биол. информация.
Модели типа (4) используют в теории биол. эволюции (включая происхождение жизни)
и в экологии. В инженерной микробиологии и теории иммунной реакции организма
используют близкие к (4) модели, но при этом учитывают зависимость коэф.
. Выбор оказывается возможным, поскольку симметричное стационарное состояние
при упомянутых условиях неустойчиво. В этом процессе возникает биол. информация.
Модели типа (4) используют в теории биол. эволюции (включая происхождение жизни)
и в экологии. В инженерной микробиологии и теории иммунной реакции организма
используют близкие к (4) модели, но при этом учитывают зависимость коэф. 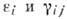 от времени.
от времени.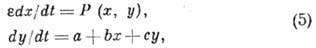
 ,
а функция P (х, у)такова, что изоклина y1(X)(решение
ур-ния Р(х, у)=0)имеет два экстремума. Модель описывает генерацию стандартного
сигнала в ответ на малое, но конечное внеш. воздействие и релаксац. автоколебания.
При изменении параметров модели (5) режим "ждущего" стационарного
состояния переходит в режим автоколебаний (и обратно). Модель (5) используют
при описании генерации нервного импульса, возникновения биол. ритмов (т. н.
биол. часов), в теории мембранной регуляции клеточного цикла и моделировании
др. явлений.
,
а функция P (х, у)такова, что изоклина y1(X)(решение
ур-ния Р(х, у)=0)имеет два экстремума. Модель описывает генерацию стандартного
сигнала в ответ на малое, но конечное внеш. воздействие и релаксац. автоколебания.
При изменении параметров модели (5) режим "ждущего" стационарного
состояния переходит в режим автоколебаний (и обратно). Модель (5) используют
при описании генерации нервного импульса, возникновения биол. ритмов (т. н.
биол. часов), в теории мембранной регуляции клеточного цикла и моделировании
др. явлений.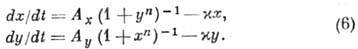
 система (6) может иметь либо одно (устойчивое) стационарное состояние, либо
три (два устойчивых и одно неустойчивое). В последнем случае система (6) при
заданном наборе параметров способна функционировать в двух разных режимах.
Переключение из одного состояния в другое возможно при изменении динамич. переменных
х и у за счёт внеш. сил (силовое переключение) и за счёт временного
изменения параметров с последующим возвращением их к исходным значениям (параметрич.
переключение). Модель (6) используют для описания дифференциации клеток при
эволюции организма и для исследования возможностей параметрич. управления онтогенезом.
система (6) может иметь либо одно (устойчивое) стационарное состояние, либо
три (два устойчивых и одно неустойчивое). В последнем случае система (6) при
заданном наборе параметров способна функционировать в двух разных режимах.
Переключение из одного состояния в другое возможно при изменении динамич. переменных
х и у за счёт внеш. сил (силовое переключение) и за счёт временного
изменения параметров с последующим возвращением их к исходным значениям (параметрич.
переключение). Модель (6) используют для описания дифференциации клеток при
эволюции организма и для исследования возможностей параметрич. управления онтогенезом.
