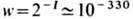 ,
число актов в биосфере за время её существования
~1030, т.е. реализация этого случая в течение времени существования
Вселенной (~1010 лет) невероятна. В этом первая проблема Э. б.
,
число актов в биосфере за время её существования
~1030, т.е. реализация этого случая в течение времени существования
Вселенной (~1010 лет) невероятна. В этом первая проблема Э. б.Эволюция биологическая - процесс развития биосферы от образования живых объектов до совр. её состояния. Выделяют след,
этапы происхождения жизни:
1) синтез осн. биологически важных молекул: Сахаров,
липидов, нуклеотидов, аминокислот и их случайных полимеров; образование комплексных
структур, содержащих полипептиды в липидной оболочке;
2) образование комплексов из полинуклеотидов
и белков, способных к комплементарной ауторепродукции (гиперциклов); образование
единого биол. кода;
3) образование многообразия живых объектов, существующих
за счёт материалов, накопленных в предбиол. период (анаэробы);
4) появление организмов, способных усваивать
энергию света, разлагать воду и использовать продукты разложения (кислород и
восстановленные органич. вещества) для биосинтеза; образование структуры трофич.
уровней, биоценозов и формирование совр. биосферы;
5) появление разумных живых существ.
Первые два этапа относят к периоду хим. (предбиол.)
эволюции; собственно Э. б. начинается с 3-го этапа.Во
всём процессе Э. б. и в каждом из периодов различают конвергентные и дивергентные
стадии. В результате конвергентной стадии свойства живых объектов становятся
одинаковыми (стадии образования единого кода и формирования вида). В дивергентной
стадии вид расщепляется, т. е. из одной популяции образуются две (или более)
различающиеся по свойствам популяции (стадии образования первичных гиперциклов,
появления многообразия живых объектов, зарождения новых видов при освоении новой
экологич. ниши, в частности новых источников питания и энергии). В процессе
дивергентной стадии численность нового вида возрастает экспоненциально, затем
рост прекращается, в конвергентной стадии численность постоянна. По исчерпании
ресурсов экологич. ниши численность резко падает и часто вид исчезает; кривая
зависимости численности от времени называется логистической. Переход от дивергентной
фазы к конвергентной совершается быстро и сопровождается большими флукту-ациями.
Матем. моделирование переходных процессов в Э. б. показывает, что часто они
носят характер катастрофы (см. Катастроф теория ),что и объясняет резкие
изломы логистич. кривой.
Свойства объектов популяции отличаются друг от
друга, т. е. имеется распределение свойств. Свойства разных популяций также
различны, в то же время их распределения могут перекрываться. Различают генотипические
и фе-нотипические свойства. Первые связаны с информацией, заключённой в генотипе
организма, т. е. с последовательностью нуклеотидов в ДНК (см. Полимеры биологические), вторые отражают феноменологию живых существ. В значит. мере фенотипич. свойства
определяются генетич. информацией (безусловные рефлексы, врождённые признаки),
но часть их связана с конкретными условиями развития и существования данного
организма или популяции (приобретённые признаки, условные рефлексы) и не связана
с генетич. информацией.
Здесь будут рассмотрены молекулярные (физ.) и
инфор-мац. аспекты Э. б. В основе этого рассмотрения лежат след. процессы, изучение
к-рых - одна из задач биофизики: а) комплементарная ауторепродукция (самовоспроизведение)
гена; б) изменение гена за счёт точечных мутаций, вызванных либо тепловыми флуктуациями,
либо мутагенными факторами (облучение УЗ и рентг. излучением, прогонами и электронами
высоких энергий, хим. мутагены и т. д.).
В результате в ДНК происходят замены нуклеотидов, их удаление и вставки; в)
блочные мутации, т. е. удаления из генома и вставки в него целой последовательности
нуклеотидов за счёт спец. аппарата клетки, позволяющего встраивать в её геном
участки ДНК (т. н. плазмиды), попадающие в клетку извне.
Во всех упомянутых случаях происходит изменение
свойств синтезируемых белков и (или) появление новых белков, что влияет на фенотипич.
признаки. Существуют механизмы регуляции на эпигенетич. уровне. Регуляция транскрипции
на уровне инициации является результатом экспрессии генома, его репрессии и
блокировки. При блокировке (спец. белками) соответствующий ген не участвует
в биосинтезе белка, не влияет на фенотипич. признаки, но участвует при редупликации
ДНК и передаётся по наследству при делении клетки. Кол-во такой нейтральной
генетич. информации (т. е. "молчащих" генов) достаточно велико.
При определ. условиях может происходить их дебло-кировка: "молчащие"
гены экспрессируются и перестают быть "нейтральными".
Регуляция транскрипции на уровне последовательности
РНК, уже скопированной с генома, происходит за счёт белков рестриктаз и лигаз,
к-рые "разделяют" РНК на блоки; нек-рые из них отбрасываются (интроны),
а другие (экзоны) сшиваются лигазами и поступают в аппарат биосинтеза белков.
Интроны можно рассматривать как нейтральную информацию, к-рая при изменении
условий (т. е. состава рестриктаз и лигаз) может превратиться в активную.
Эти процессы играют разную роль в конвергентных
и дивергентных стадиях Э. б. В конвергентных стадиях гл. факторы - естеств.
отбор (принцип Дарвина) и выбор. Отбор происходит по фенотипич. признакам, и
в результате него в живых остаются наиб. приспособленные к данной экологич.
нише особи. Отбор влияет на геном, именно на экспрессированную его часть, при
этом вымирают те особи, в к-рых совокупность белков (синтезированных на основании
генетич. информации) не обеспечивает наиб. приспособленного фенотипа. В результате
распределения по фенотипич. и генетич. свойствам сужаются.
Отбор протекает на фоне точечных мутаций. Те
из них, к-рые снижают эволюц. преимущества, элиминируются (вымирают). Малая
часть мутаций, дающих эволюц. преимущества, сохраняется и закрепляется. Блочные
мутации, ведущие к сильному изменению фенотипич. признаков, в конвергентной
стадии не участвуют (или элиминируются как негативные).
Выбор означает вытеснение одного вида другим,
даже если они фенотипически равноправны, но взаимодействие между ними антагонистично
(межвидовая борьба сильнее, чем внутривидовая, и перекрёстное оплодотворение
исключено). В результате выбора выживает более много-числ. вид, не всегда наиб.
приспособленный. Выбор играет важную роль на ранних этапах эволюции - при образовании
генетич. кода и возникновении биол. асимметрии, а также при освоении одной экологич.
ниши двумя видами. При этом один вид вытесняется другим, так что одну нишу может
заселять лишь один вид. Мутации в процессе выбора существ. роли не играют.
Гл. фактор дивергентных стадий - возникновение
новых фенотипич. свойств, позволяющих начать освоение новой ниши. Они могут
появиться в результате либо блочных мутаций, либо перераспределения экзонов
и нитронов, либо деблокировки "молчащего" генома.
1. Возникновение жизни
Молекулярный аспект. Накопление биологически
важных молекул (липиды, сахара, аминокислоты, нуклеотиды) в необходимых для
Э. б. кол-вах в термодинамически равновесных условиях невозможно. В открытой
системе оно возможно благодаря воздействию УФ и космич. излучения, электрич.
разрядам и перепадам температуры. Наиб, вероятно образование и накопление упомянутых
молекул в горячих водоёмах в результате вулканич. деятельности.
В тех же условиях возможны спонтанная полимеризация
нуклеотидов и белков и самоорганизация их в капли, покрытые липидной мембраной
(коацерваты, мариграны, микросферы). Однако комплементарная ауторепродукция
ДНК в этих условиях практически невозможна.
Информационный аспект. В совр. биосфере
репродукция ДНК осуществляется белком - репликазой. Биосинтез репликазы происходит
на основе информации, содержащейся в соответствующем гене (участке ДНК). Эта
информация записана в ДНК в форме последовательности из четырёх разных нуклеотидов.
Трансляция её в последовательность аминокислот (20-буквенный код) происходит
на основании кода, согласно к-рому каждой тройке нуклеотидов (кодону) соответствует
определ. аминокислота. Этот код (т. е. алгоритм соответствия) одинаков во всей
биосфере - от простейших организмов до человека. В совр. коде каждому аминокислотному
остатку соответствует неск. кодонов, т. е. код вырожден. Однако обратное вырождение,
т. е. случаи, в к-рых одному кодону соответствует неск. разных аминокислот,
практически отсутствует (имеются редкие исключения: в генетич. материале митохондрий,
представляющих собой автономные внутриклеточные органеллы). Аппарат биосинтеза
содержит набор адаптеров (белки - аминоациладенилат синтетазы), несущий информацию
о коде, и рибосомы, осуществляющие синтез белковой последовательности при участии
реплики РНК и транспортных РНК, ответственных за связь между рибосомой и адаптерами.
T. о., имеется замкнутый цикл: белки катализируют ауторепродукцию ДНК, геном
регулирует синтез этих белков. Система, содержащая такой цикл, названа гиперциклом.
Кол-во необходимой информации, содержащейся в
аппарате биосинтеза, достаточно велико (I~ 1000 бит). Вероятность случайного
возникновения такой системы за один акт,
число актов в биосфере за время её существования
~1030, т.е. реализация этого случая в течение времени существования
Вселенной (~1010 лет) невероятна. В этом первая проблема Э. б.
Предлагаемое решение проблемы сводится к следующему.
Первичный гиперцикл содержит всего две компоненты: двойную спираль (биспираль)
ДНК произвольной последовательности и белок, имеющий форму чехла, окружающего
биспираль, в периодически изменяющихся условиях (температуры, растворимости и т.
д.) функционирующий как репликаза. Синтез такого белка может происходить путём
полимеризации аминокислот, предварительно адсорбированных на биспирали ДНК (полимеризация
на гетерогенном катализаторе), образующийся полимер - слепок с поверхности катализатора.
При адсорбции аминокислот на биспирали ДНК возможно
избират. связывание аминокислот с определ. кодо-нами, имеющими к ним хим. сродство.
Благодаря этому при последующей полимеризации образуется белок-чехол, аминокислотная
последовательность к-рого коррелирует с последовательностью кодонов в матричной
ДНК. Вероятность такого процесса не мала, поскольку он протекает без участия
кода, трансляции, аппарата биосинтеза и не нуждается в содержащейся в них информации.
След. проблема - образование адаптера, т. е.
молекул, комплементарных, с одной стороны, к кодону, а с другой- к аминокислотному
остатку. Синтез их также может происходить на основе гетерогенного катализа.
Набор адаптеров в случае отсутствия обратного вырождения (т. е. адаптеров, комплементарных
к одинаковым кодо-нам и разным аминокислотам) существенно ускоряет репродукцию
гиперцикла; наличие же обратного вырождения ведёт к прекращению репродукции
и гибели гиперциклов.
Совокупность одинаковых гиперциклов, образовавшихся
в результате комплементарной авторепродукции одной биспирали ДНК с одинаковым
набором адаптеров, представляет собой первичный живой объект, обладающий информацией
о коде. Популяция с др. предшественником (биспирали ДНК с др. последовательностью)
обладает, вообще говоря, др. набором адаптеров, поскольку избирательность
взаимодействия аминокислот и кодонов не столь велика, чтобы обеспечить однозначность
алгоритма соответствия. Поэтому множество объектов с разными предшественниками
ещё не представляет собой первичную биосферу, поскольку каждый из объектов обладает
своим вариантом кода.
Проблема образования единого кода сводится к
проблеме выбора в условиях сильного антагонистич. взаимодействия разных объектов,
когда их адаптеры смешиваются и возникает обратное вырождение. Выбор единого
кода описывается матем. моделью типа Вольтера - Лотки:
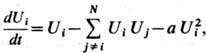
где 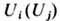 -концентрация
объектов i-го ( j-гo ) типа, t - время; первый член описывает репродукцию, второй- антагонистич. взаимодействие
разных объектов, третий- эффект "тесноты", т. е. конкуренцию одинаковых
объектов. В случае, когда внутривидовой антагонизм меньше межвидового (а<1),
модель описывает образование чистого состояния, где остаётся одна из N
возможных популяций (напр., i-тая), а все остальные вымирают:
-концентрация
объектов i-го ( j-гo ) типа, t - время; первый член описывает репродукцию, второй- антагонистич. взаимодействие
разных объектов, третий- эффект "тесноты", т. е. конкуренцию одинаковых
объектов. В случае, когда внутривидовой антагонизм меньше межвидового (а<1),
модель описывает образование чистого состояния, где остаётся одна из N
возможных популяций (напр., i-тая), а все остальные вымирают: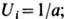
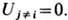 Симметричное состояние, где все
Симметричное состояние, где все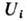 одинаковы,
неустойчиво; какая из популяций победит, зависит от нач. концентраций и случайных
флуктуации. Выбор единого кода представляет собой конвергентную стадию, в результате
к-рой формируется биосфера, однако ещё без многообразия её совр. состояния.
одинаковы,
неустойчиво; какая из популяций победит, зависит от нач. концентраций и случайных
флуктуации. Выбор единого кода представляет собой конвергентную стадию, в результате
к-рой формируется биосфера, однако ещё без многообразия её совр. состояния.
Проблема возникновения биол. асимметрии решается
аналогично. В совр. живой природе оптически активные вещества (сахара и аминокислоты)
присутствуют не в ра-цемич. смеси: преобладают правовращающие сахара и ле-вовращающие
аминокислоты. Рацемич. смеси невыгодны, т. к. в этом случае необходимо иметь
удвоенный набор белков-ферментов - отдельно для левых и правых изомеров. Поэтому
в живой природе могут существовать чистые ("левые" или "правые")
популяции, поскольку эти варианты равноправны. Взаимодействие между разл. популяциями
антагонистично - "правые" сахара являются ядом для "левых"
объектов и наоборот.
2. Проблема биологического разнообразия (дивергентная
стадия)
По мере исчерпания ресурсов возникает необходимость
появления белков-ферментов с новой функцией, способных утилизировать новые ресурсы.
Расщепление популяции (дивергенция) происходит в результате образования нескольких
новых белков с разными функциями, что позволяет осваивать разные биол. ниши. При
этом необходимо сохранить белки, выполняющие прежние функции.
После выбора единого кода и возникновения аппарата
трансляции образуются белки с новой функцией, т. е. появляются новые участки ДНК,
кодирующие эти белки. Это реализуется в процессах увеличения генома путём накопления
тождественных копий и последующего накопления мутаций до тех пор, пока одна
из копий не приобретет нужные свойства.
функции белка-фермента определяются строением активного
центра (специфич. функции) и структурой глобулы. От последней зависят стабильность
белка, его способность к конформац. изменениям, растворимость в воде и (или)
возможность внедрения в липидные мембраны (см. Клеточные структуры, Растворы). Новые специфич. функции возникают за счёт точечных мутаций участка ДНК, кодирующего
активный центр, а новые структурные функции - за счёт блочных мутаций - т. е.
новой комбинации участков ДНК. Кол-во возникающей новой ценной информации, необходимое
для синтеза функционального белка, оценивается в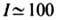 бит,
осн. её часть (~ 70-80 бит) связана со структурными изменениями, сравнительно
небольшая часть (~20-30 бит) необходима для организации активного центра, ответственного
за специфику функции. Вероятность спонтанного образования объекта, содержащего
информацию I~ 100 бит за один акт,
бит,
осн. её часть (~ 70-80 бит) связана со структурными изменениями, сравнительно
небольшая часть (~20-30 бит) необходима для организации активного центра, ответственного
за специфику функции. Вероятность спонтанного образования объекта, содержащего
информацию I~ 100 бит за один акт,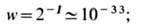
т. о., вероятность возникновения даже одного нового
белка оказывается малой.
В Э. б. известны случаи, когда новая функция, дающая
большие преимущества, связана с появлением сразу нескольких новых белков. При
этом образование каждого нового белка в отдельности не даёт эволюц. преимуществ.
Такие "большие скачки" эволюции наз. ароморфозами; примером является
возникновение аппарата фотосинтеза .В этом случае кол-во информации порядка
сотен бит и вероятность её возникновения абсурдно мала. Время, необходимое для
реализации рассматриваемых процессов Э. б., значительно превышает время жизни
Вселенной.
Существуют пути преодоления этих трудностей теории.
Так, за счёт блочных мутаций значит, часть структурной информации может быть
заимствована из уже имеющихся генов, а не создаваться заново. Новая структурная
информация возникает в результате перестановки небольшого числа уже существующих
блоков, в к-рых уже содержится нужная информация. Для этого необходим достаточный
запас нейтральной информации в форме "молчащих" генов и нитронов.
В этом случае кол-во действительно новой информации значительно снижается и
вероятность её возникновения возрастает. (Аналогично в техн. эволюции - при
создании новой машины конструктор использует уже существующие стандартные детали,
а не изобретает их заново, новой является лишь комбинация деталей.)
Рассмотрим примеры получения новой структурной
информации.
1. Формирование единого кода протекает на фоне
избытка субстратов, синтезированных в предбиол. период. После их исчерпания
накапливаются остатки отмерших организмов. Для их утилизации необходимы белки,
расщепляющие полинуклеотиды (нуклеазы) и полипептиды (протеазы), структура к-рых
сходна - глобулы с ложбиной. Такие белки могут быть получены из первичного белка
- репликазы. Для этого достаточно "разрезать" молекулу вдоль оси
и в разрез "поместить" активные гидролизующие аминокислоты (серии,
гистидин и т. п.). Набор последних различен в нуклеазах и протеазах, что и определяет
их специфичность. Слова "разрезать" и "поместить" означают,
что в запасной копии гена, кодирующего репликазу, заменяются неск. кодонов (за
счёт точечных мутаций) так, что образующийся белок приобретает упомянутые свойства.
Затем этот ген выделяется и экспрессируется (блочная мутация).
2. Для образования белков аппарата фотосинтеза
необходимы мембранные белки, способные поглощать свет. Мембранные белки содержат
большое кол-во структурной информации: на боковых поверхностях молекулы белка
расположены гидрофобные аминокислотные остатки, а на торцевых - гидрофильные.
Напр., молекула бактериоро-допсина состоит из семи столбов (a-спиральных
участков). Каждый столб содержит ок. 30 аминокислот, в к-рых чередуются две
гидрофобные и две произвольные. Благодаря этому половина поверхности a-спирального
столба оказывается сплошь гидрофобной; именно такие столбы образуют боковую
поверхность бактериородопсина.
Необходимое для этого процесса кол-во информации
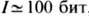 , и вероятность её возникновения мала. Вероятность же возникновения одного столба
(I~ 15 бит) в течение времени ~1010 лет не мала, и процесс
образования гена, кодирующего один столб, вполне вероятен. Накопление семи копий
этого гена и соединение их в цепь (за счёт блочных мутаций), кодирующую весь
мембранный белок, тоже достаточно вероятно.
, и вероятность её возникновения мала. Вероятность же возникновения одного столба
(I~ 15 бит) в течение времени ~1010 лет не мала, и процесс
образования гена, кодирующего один столб, вполне вероятен. Накопление семи копий
этого гена и соединение их в цепь (за счёт блочных мутаций), кодирующую весь
мембранный белок, тоже достаточно вероятно.
3. Согласно принципу преемственности, возникновение
новых функций не должно мешать выполнению и сохранению прежних. В сложных организмах
эти условия дополнительны (дуальны). Так, воздействие мутагенных факторов полезно
для новых функций и вредно для прежних. Проблема дуальности была решена путём
удвоения всего генетич. материала (за счёт блочной мутации) и появления перекрёстного
полового размножения. При этом отбор особей женского пола происходил по фенотипич.
признакам (анатомическим и физиологическим), способствующим защите репродуктивных
органов и сохранению генома. Напротив, отбор мужских особей происходил по признакам,
способствующим накоплению новой информации.
4. Обмен генетич. информацией между клетками
увеличивает фонд запасённых генов, что полезно в дивергентных стадиях. Он может
происходить: а) в результате внедрения участков генома (плазмид) одной клетки
в другую и встраивания его в ДНК; такой процесс происходит в колониях одноклеточных
организмов, при внедрении чужеродной клетки в организм и между разными клетками
одного организма; б) в результате внедрения одной клетки в другую и образования
симбиотич. организма (напр., хлоропластов растит. клеток и митохондрий); в)
при скрещивании; в этом случае увеличение информации происходит не путём увеличения
ДНК, а благодаря удвоению числа хромосом.
3. Дискуссионные вопросы теории биологической
эволюции
1) Согласно теории естеств. отбора, любая нейтральная
генетич. информация должна элиминироваться как лишняя нагрузка, не дающая эволюц.
преимуществ. Согласно нейтралистской теории эволюции, нейтральная информация
должна сохраняться как материал, необходимый для выбора подходящего варианта
при дивергенции. Разрешение этого противоречия состоит в том, что Э. б. представляет
собой чередование конвергентных и дивергентных стадий. При конвергенции часть
информации действительно элиминируется как избыточная, но значит. часть её остаётся
на случай перехода в дивергентную фазу.
2) Согласно ортодоксальному дарвинизму, благоприятные
фенотипич. признаки, приобретённые в результате упражнений, но не в результате
мутаций, не могут передаваться потомству. Однако противоположное утверждение
Ж. Б. Ламарка (J. В. Lamarck) не исключал и Ч. Дарвин (С. Darvin). Молекулярной
основой первого утверждения является догма Крика, согласно к-рой белки (и др.
метаболиты) не могут влиять на геном. Однако известные ныне и перечисленные
выше механизмы генной регуляции допускают нарушение догмы Крика из-за влияния
белков на эпигенетич. процессы, образование плазмид и внедрение их в геном репродуктивных
органов. Важность разрешения этой проблемы в том, что даже редкие случаи наследования
благоприобретённых признаков существенно ускоряют эволюц. процесс.
3) Ряд заболеваний связан с тем, что в клетку
внедряется патогенный вирус, размножается в ней (активный вирус) и (или) встраивается
в геном и размножается при его редупликации (провирус). В клетках организма
обнаружен аппарат, способствующий этим процессам. Возникает вопрос: каким образом
в Э. б. возник и сохранился аппарат, явно не способствующий выживанию данного
организма?
Возможное решение вопроса состоит в том, что геном вируса аналогичен плазмиде и аппарат, приспособленный для обмена генетич. информацией, способствует развитию вирусных заболеваний. Последнее - плата организма за ускорение Э. б., в результате к-рой этот организм образовался.

|
|
