Биологические полимеры (биополимеры) - природные макромолекулы, играющие осн. роль в биол. процессах. К П. б. относятся
белки, нуклеиновые кислоты (НК) и полисахариды. П. б. образуют структурную основу
всех живых организмов; все процессы в клетке связаны с взаимодействиями П. б.
между собой и с др. молекулами. Среди последних
важную роль играют л и p и д ы, образующие биол. мембраны (см. Клеточные
структуры). Липиды не являются полимерами, но обладают нек-рыми общими с
ними свойствами, в частности способностью образовывать жидкокристал-лич. структуры.
Биологические полимеры являются высокомолекулярными соединениями
(мол. массаа.
е. м)., к ним приложимы все закономерности, установленные для др. природных
и синтетич. полимеров. Однако особенности хим. строения приводят к появлению
у П. б. уникальной пространств, структуры, необычных физ., хим. и биол. свойств.
По строению осн. цепи белки и НК однородны, подобно г о-мополимерам, у к-рых
все мономерные звенья цепи идентичны. Но в последовательности боковых групп
у П. б. закодирована генетич. информация организма, поэтому П. б. следует отнести
к гетеропо-лимерам с заданной нерегулярной последователь-ностью мономерных звеньев.
В структуре и свойствах П. б. отражены эти особенности их хим. строения. Пространств.
строение П. б. с определ. структурой всей макромолекулы наз. конформацией; от
конформации зависит взаимодействие П. б. с др. молекулами. Наиб. важные биол.
функции П. б. также определяются его конформацией и способностью изменять её при
разл. взаимодействиях. В большинстве случаев взаимодействия П. б. являются специфически-м
и, т. е. зависят от последовательности мономерных звеньев и локальной структуры
(см. также Биофизика).
Различают 4 уровня структурной организации П.
б. Наиб. отчётливо они выражены у белков. Первичная структура - это хим. строение
молекулы. Чаще всего под первичной структурой понимают последовательность мономерных
звеньев П. б. В первичную структуру включаются хим. связи между цепями и внутри
цепей (между отд. звеньями). Вторичная структура - спиральное расположение мономерных
звеньев в тех или иных участках цепи П. б. Третичная структуrа - пространств.
структура цепи, включая расположение элементов вторичной структуры и связывающих
их участков. Четвертичная структура - расположение отд. цепей (единиц третичной
структуры) в образуемом ими комплексе.
Белки состоят из одной или неск. полипептидных
цепей, к-рые соединены между собой хим. или межмолекулярными связями. Полипептидные
цепи построены из мономерных звеньев - аминокислотных остатков 20 разл. сортов.
Аминокислоты представляют собой органич. (карбоновые) кислоты, содержащие 1
или 2 аминогруппы NH2. В нейтральной среде они имеют структуру,
где R - боковая группа, своя для каждой из 20
аминокислот. Аминокислоты являются оптич. L-изомерами (см. Изомерия молекул). Число мономерных звеньев, входящих в полипептидные цепи, может изменяться
от неск. десятков до неск. тысяч; полипептиды с меньшим числом звеньев наз.
олигопепти-д а м и. Каждый белок имеет определ. размеры (мол. масса
его индивидуальность определяется последовательностью
аминокислотных остатков. По своим функциям белки делятся на каталитические (ферменты,
биол. катализаторы хим. реакций), структурные, транспортные (гемоглобин), рецепторные,
ре-гуляторные (гормоны), защитные (антитела) и др. В зависимости от состава
выделяют простые белки- протеины, состоящие только из аминокислот, и сложные
белки -протеиды, в состав к-рых наряду с аминокислотами входят углеводы (гликопротеиды),
липида (липопротеиды), НК (нуклеопротеиды) и т.
д. По форме различают глобулярные белки, образующие плотные глобулы, и фибриллярные
белки, образующие длинные волокна или слои. Белки участвуют в важнейших генетич.
и регуляторных процессах. Нек-рые структурные белки могут образовывать агрегаты
в виде волокон, трубочек, оболочек. Иногда один и тот же белок выполняет неск.
функций.
Первичная структура. Образование поли-пептидной
цепи с заданной последовательностью аминокислотных остатков происходит в клетке
внутри клеточного аппарата - рибосомы. Присоединение каждого последующего звена
цепи происходит с выделением молекулы воды. Образующаяся цепь имеет следующую
структуру:
поскольку соединение мономеров происходит по
принципу "голова к хвосту", цепь определ. образом направлена: слева
находится N-конец цепи, справа - С-конец. Аминокислотные остатки цепи в зависимости
от вида боковой группы R делятся на неск. типов. К неполярным, плохо растворяющимся
в воде относятся аланин, валин, лейцин, изолейцин, фенилаланин, триптофан, тирозин,
метионин, глицин и цистепин. Полярные и заряженные аминокислотные остатки обладают
хорошей растворимостью в воде. К полярным относятся серии, треонин, аспарагин,
пролин и глутамин. Заряжены аспарагиновая и глутаминовая к-ты (отрицательно),
лизин и аргинин (положительно). Могут быть заряженными также цистеин и гистидин.
В целом молекула белка несёт положит. и отрицат. заряды. В первичной структуре
белка заключена вся информация, определяющая его пространств. структуру и функции.
Определение первичной структуры полипептидной цепи производят путём частичного
расщепления её на короткие перекрывающиеся фрагменты с последующим анализом
их аминокислотной последовательности, начиная с N-конца. Это удаётся сделать
для не слишком длинных последовательностей, поэтому структуру длинных полипептидов
находят, комбинируя данные для фрагментов. Полипептидная цепь обладает гибкостью
за счёт вращения вокруг хим. связей, образуемых атомами С (чёрные шарики на
рис. 1, вращение изображено стрелками). Связь между группами СО и NH
наз. пептидной. Вращение вокруг пептидной связи затруднено, поэтому атомы H,
N, С и О лежат в одной плоскости. Вторичная структура. Благодаря своей
гибкости полипептидные цепи способны образовывать упорядоченные структуры со
спиральной симметрией. Наиболее распространеныспирали
и структуры. a-Спираль
представляет собой правую спираль, у к-рой на один виток приходится 3,6 аминокислотных
остатка; шаг спирали 5,4 ,
диаметр(без боковых
групп).
Рис. 1. Вращение пептидных групп.
Спираль стабилизирована водородными связями
между группами СО и NH разл. мономерных звеньев, отстоящих друг от
друга на расстоянии 4 остатков. Водородные связи (пунктир на рис. 2,а)
направлены вдоль оси спирали, в целомспираль
представляет собой довольно жёсткую структуру. Не всем аминокислотным остаткам
энергетически выгодно образование a -
спирали. Знание соответствующих энергетич. параметров позволяет предсказывать
вероятность образования a-
спирали в том или ином участке белка. Существуют b-слои двух типов: параллельные
и антипарал-лельные. На рис. 2,б показана структура антипарал-лельногослоя.
Стабилизирующиеслой
водородные связи между пептидными группами
направлены поперёк цепей, а сами цепи вытянуты и образуют складчатую структуру.
В белке встречаются также т. н.
Рис. 2. Вторичная структура белков: а
- a-спираль; б - b-структура.
b- изгибы, обеспечивающие поворот цепи примерно
на 180 при образовании
водородной связи. Возможны и др. типы спиралей. Все названные вторичные структуры
характерны для глобулярных белков. Фибриллярный белок, из к-рого строятся длинные
ориентиров. волокна, образует спирали иного вида. Вторичную (и третичную) структуру
белка исследуют с помощью рентгеновского структурного анализа, позволяющего
определить положение всех атомов в молекуле. Трудности здесь связаны с тем,
что не каждый белок можно получить в виде кристаллов необходимого размера. Обычно
структура белка в растворе мало отличается от структуры в кристалле, это связано
с тем, что кристаллы белка содержат много воды. Однако в целом вопрос о соответствии
структуры белка в растворе и в кристалле остаётся открытым. Содержаниеa-
и b-структур
сильно различается для разл. белков.
Третичная структура. Большинство глобулярных
белков находится в водно-солевой среде. Укладка элементов вторичной структуры
при этом такова, что гидрофильные (полярные, заряженные) аминокислотные остатки
располагаются в осн. на поверхности глобулы, а неполярные, плохо растворимые
в воде (гидрофобные) аминокислотные остатки - во внутр. части глобулы. При этом
глобула приобретает уникальную (идентичную для всех молекул данного белка) компактную
и стабильную форму.
Чаще всего внутр. часть глобулы образована b-слоя-ми,
а наружная -спиралями.
Установлена закономерность в аминокислотной последовательности в этихспиралях:
каждое 3-е или 4-е положение вдоль цепи занимают неполярные аминокислотные остатки.
При этом на боковой поверхности цилиндра, к-рым можно представить спираль,
образуется неполярная полоса, параллельная её оси. Именно эта гидрофобная полоса
обращена внутрь глобулы и контактирует с её гидрофобной частью.
Исключение составляют мембранные белки, контактирующие
с неполярной жирной внутр. частью липидной мембраны. На поверхности белка в
этом случае находятся гидрофобные аминокислотные остатки.
Ещё одна важная закономерность пространств. структуры
белков - доменное строение. Часто единая поли-пептидная цепь образует не одну
глобулу, а неск. компактных областей, расположенных определ. образом в пространстве.
Каждая такая область (домен) формируется из спиралей,
слоев и др.
элементов вторичной структуры. В этом случае можно говорить как о третичной
структуре таких доменов, так и о третичной структуре белков в целом, понимая
под этим взаимное расположение доменов в пространстве. Примером домена, содержащегося
во мн. белках, является блок из двухслоев,
соединённых между собойспиральным
сегментом. Доменная структура белков важна для их биол. функций. Вероятно также,
что домены - это элементарные белки, на основе к-рых в ходе эволюции возникает
разнообразие белковых структур.
Четвертичная структура. В тех случаях, когда
глобулярный белок состоит из неск. субъединиц, не связанных между собой хим.
связями, говорят о его четвертичной структуре. Связь субъединиц между собой
осуществляется гл. обр. за счёт гидрофобных взаимодействий; при этом на контактирующих
частях поверхности субъединиц расположены в осн. гидрофобные аминокислотные
остатки. Иногда во взаимодействие между субъединицами глобулярных белков дают
заметный вклад водородные связи. Др. тип четвертичных структур представляют
белки, образующие нити цитоскелета. Цитоскелет заполняет пространство между
ядром и внутр. поверхностью клеточной мембраны и выполняет ряд важных функций,
определяя форму клетки, её перемещение как целого, размещение и транспорт внутр.
компонентов. Известны три типа таких нитей: микрофиламенты, микротрубочки и
промежуточные филаменты. Подробно изучены первые два типа. Микрофиламенты собираются
из молекул глобулярного белка актина, соединяясь в длинные цепи, образующие
двойные спирали. Микротрубочки также собираются из глобулярных молекул белка
тубулина и являются важным компонентом ми-тотич. аппарата (аппарата деления)
клетки, образующим т. н. митотич. веретно и определяющим распределение генетич.
материала между дочерними клетками.
Особый тип структур представляют фибриллярные
белки актин и миозин, образующие упорядоченные структуры (саркомеры). Их скольжение
друг относительно друга составляет основу механизма мышечного сокращения. В
сложные пространств. структуры собираются белки оболочек вирусов, бактериофагов
и таких структур, как рибосомы, нуклеосомы и др.
Высшие структуры белков - это состояния, обладающие
относит. минимумом свободной энергии. Они устойчивы в физиологич. условиях,
могут изменяться лишь в определ. пределах. Наиб. устойчива первичная структура
белков, остальные легко разрушаются при внеш. воздействиях. Такое разрушение
наз. денатурацией и, как правило, приводит к потере биол. свойств.
Нуклеиновые кислоты. Дезоксирибонуклеиновые кислоты
(ДНК) и рибонуклеиновые кислоты (РНК) являются полинуклеотидами, т. е. П.б.,
мономерными звеньями к-рых служат нуклеотиды. Нуклеотиды
состоят из азотистого основания, остатков фосфорной к-ты и углевода (рибозы
или дезокси-рибозы). ДНК является хранителем генетич. информации организма,
записанной в виде последовательности 4 сортов её мономерных звеньев. Эта информация
переписывается (транскрибируется) при синтезе информац. (матричной) РНК (мРНК),
а затем с помощью генетич. кода переводится (транслируется) в аминокислотную
последовательность белков. Др. виды РНК выполняют роль переносчиков аминокислот
(транспортные РНК - тРНК) или составляют структурную основу рибосом (рибосомные
РНК - рРНК). Молекулы РНК в нек-рых случаях могут обладать также каталитич.
активностью, подобной активности белков-ферментов (т. н. рибозимы).
Первичная структура НК. Полинуклео-тидная цепь
(рис. 3) состоит из сахарофосфатного остова (в него входит дезоксирибоза в случае
ДНК и рибоза в случае РНК), к к-рому присоединены плоские боковые группы - азотистые
основания (аденин А, цитозин С, гуанин G и тимин Т в случае
ДНК; А, С, G и урацил U в случае РНК). В клетке такие цепи синтезируются
с помощью спец. ферментов на матрице- молекулах
ДНК; существует и процесс синтеза ДНК на РНК-матрице, осуществляемый др. ферментом
(обратной транскриптазой). Полинуклео-тидная цепь имеет направление, определяемое
тем, что -и
атом С одного мономера соединяется фос-фодиэфирной связью с -м
атомом С следующего мономера. Каждая мономерная группа цепи ионизована p
несёт один отрицат. заряд. Размеры молекул РНК и ДНК изменяются в широких пределах.
Транспортные РНК (самые короткие молекулы РНК) состоят из 75-84 нуклеотидов;
длина гетерогенных ядерных РНК достигает
нуклеотидов. Короткие ДНК содержат обычно
неск. тысяч пар нуклеотидов, но существуют ДНК, к-рые содержат их
Вторичная структура ДНК. Осн. принцип образования
вторичных структур полинуклеотидов - т. н. комплементарное спаривание азотистых
оснований.
Оно приводит к образованию двойных и тройных
винтовых структур (спиралей), стабилизируемых водородными связями между азотистыми
основаниями разных цепей н межплоскостными взаимодействиями азотистых оснований.
Осн. вторичная структура ДНК (B-форма), представляющая собой правую двойную
спираль, предложена в 1953 Дж. Уотсоном (J. Watson) и Ф. Криком (F. Crick).
В этой структуре две комплементарные цепочки антипараллельны. Против каждого
А одной цепи расположен Т другой, против G расположен С (в дву-нитевой РНК А спаривается с U). При этом образуются
энергетически выгодные водородные связи: 2 в AT-паре и 3 в GC-парe; расстояние
между точками присоединения оснований к сахарам оказывается одинаковым для А
Т- и GC-паp (рис. 4). Сахарофосфатные цепи образуют при этом гладкие винтовые
линии. Плоскости оснований в 5-форме ДНК составляют с осью двойной спирали прямой
угол. На виток двойной спирали приходится в натриевой соли ДНК при высокой влажности
10 пар оснований. Расстояние между плоскостями соседних пар оснований составляет
3,4 что оптимально
для межплоскостных взаимодействий, вносящих наиб. энергетич. вклад в стабильность
двойной спирали. В растворе на виток двойной спирали в
B-форме приходится 10,5 пары оснований. Диаметр двойной спирали равен примерно
22. В-форма
характерна для натриевой соли ДНК. При изменении внеш. условий (температуры, ионного
состава среды) параметры двойной спирали в В-форме изменяются, поэтому
следует говорить о 5-семействе структур. К этому семейству относится и литиевая
соль ДНК, т. н. С-форма, в к-рой на виток двойной спирали приходится 9,3 пары
оснований, плоскость оснований отклонена на от
плоскости, перпендикулярной к оси спирали.
В натриевой соли ДНК при относит. влажности ниже
75% происходит кооперативный резкий переход ДНК из В- в Л-форму. Л-форма
(точнее А - семейство форм) - это также правая двойная спираль, но с
др. параметрами, чем у B-формы. Плоскости оснований сильно отклонены от плоскости,
перпендикулярной к оси спирали, а сами пары комплементарных оснований смещены
от оси двойной спирали к её периферии, поэтому при наблюдении вдоль оси молекула
в А -форме представляется полой трубкой. РНК существует только в A-форме,
как и гибриды ДНК - РНК. Характерная для двунитевой РНК структура содержит 11
пар оснований на виток двойной спирали, а отклонение плоскости оснований от
плоскости, перпендикулярной к осп, составляет
B-форма - осн. структура ДНК в живой клетке. ДНК может существовать и в др.
форме, в виде Z-спирали. Рентге-ноструктурный анализ позволил, как и в случае
белков, установить с высоким разрешением пространств.
структуры полинуклеотидов с разл. последовательностями нуклеотидов. Z-форма
ДНК, получившая своё назв. в связи с зигзагообразным строением сахарофосфатного
остова, представляет собой левую двойную спираль с периодом
содержащую 12 пар оснований на виток и образованную антипараллельными полинуклеотидными
цепями, спаренными по правилам комплементарности. Повторяющимся звеном в ней
является не одна пара нуклеотидов, а две. Наиб. легко в Z-форму переходят регулярно
чередующиеся последовательности пурино-вых и пиримидиновых нуклеотидов. В физиологич.
условиях Z-форма в линейных ДНК не наблюдалась. Однако в кольцевых молекулах
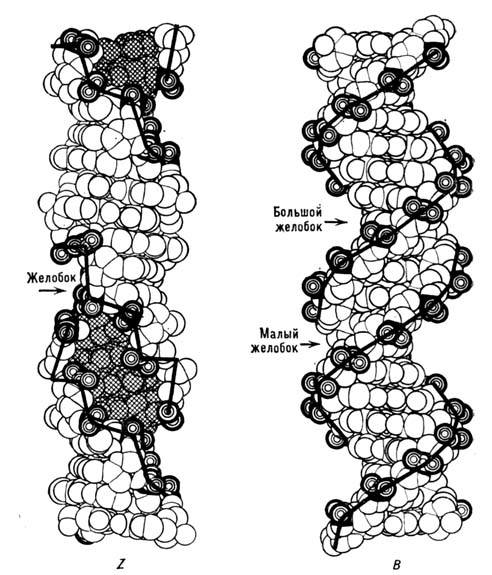
ДНК может происходить переход отд. участков молекулы в Z-форму. На рис. 5 приведены
объёмные модели ДНК в В- и Z-формах.
Рис. 4. Уотсон-криковские пары оснований (жирным
пунктиром обозначены водородные связи).
Двойная спираль ДНК в B-форме является сравнительно
жёсткой молекулой. Её макромолекулярные свойства в растворе хорошо описываются
моделью гибкого упругого стержня, совершающего тепловое движение. Изгибная жёсткость
ДНК в А-форме больше, чем в В-форме, причем она анизотропна: молекула в каждой
точке легче изгибается в направлении желобов двойной спирали, чем в перпендикулярном
направлении.
Вторичная и третичная структуры РНК. Молекулы
РНК встречаются преим. в виде одиночных нитей, в к-рых образуются двунитевые
шпильки за счёт спаривания оснований комплементарных участков нити. Однонитевые
участки могут образовывать водородные связи с др. однонитевыми участками, определяя
третичную структуру молекулы. Третичная структура хорошо изучена для молекулы
тРНК; если по вторичной структуре тРНК напоминает клеверный лист, то в пространстве
она принимает форму буквы Г. Вторичная структура фенилаланиновой тРНК, близкая
к A-форме, содержит 20 пар оснований, между к-рыми образованы 52 водородные
связи. Третичная структура содержит ещё неск. дес. таких связей с участием азотистых
оснований и сахарофосфатных цепей. Все виды тРНК имеют сходную третичную структуру.
Третичная структура ДНК. В вирусных частицах ДНК компактно упакована, однако
данные о виде этой упаковки отсутствуют. Лучше известна упаковка ДНК в хромосомах
эукариотич. клеток (см. Клеточные структуры). ДНК вирусов, бактериофагов,
плазмид и бактерий обычно представляют собой кольца, образованные замкнутыми
двойными спиралями (каждая из нитей замкнута на себя). Хромосомная ДНК в эукариотич.
клетках также образует петли, топологически эквивалентные замкнутым кольцам.
Кольцевая ДНК обычно сверхспирализована и образует пространств. сверхвитки,
к-рые также можно рассматривать как элементы третичной структуры ДНК. В разл.
условиях и в зависимости от последовательности нуклеотидов, ДНК может образовывать
и др. виды вторичной и третичной структур (параллельные спирали, тройные и четвертные
спирали и др.).
Рис. 5. Объёмные модели ДНК в В- и Z-формах (жирной
линией обозначен сахарофосфатный остов).
Полисахариды являются П. б., построенными из
мо-носахаридных остатков. Примерами линейных гомо-полисахаридов являются амилоза
(составная часть крахмала) и целлюлоза (осн. часть древесины). Мономером амилозы
и целлюлозы является глюкоза. Др. пример линейного гомополисахарида - хитин,
из к-рого построены панцири насекомых. Мономерным звеном линейного полисахарида
может быть и дисахарид. Первичная структура полисахарида, как правило, регулярна,
но существуют полисахариды с нерегулярной последовательностью разл. мономерных
звеньев. Помимо линейных существуют полисахариды с разветвлённой первичной структурой.
Линейные полисахариды образуют жёсткие вторичные структуры (одно-, двух- и трёх-нитевые
спирали). Более высокие структуры могут быть как волокнистыми, так и гелеобразными.
Если однородность полисахаридной цепи нарушена встраиванием др. сахаридов или
ветвлениями, полисахариды могут образовывать гибкие волокна или гели. Полисахариды
могут образовывать комплексы с белками и липидами, они придают жёсткость и прочность
стенкам клеток растений и бактерий. Стенки животной клетки не обладают этими
св-вами и содержат в клеточной мембране лишь нек-рое кол-во олигосахаридов (коротких
полисахаридов), связанных с белками, т. н. глико-протеидов.
Исследование структуры и свойств П. б. производят
разл. физ. и физ--хим. методами. Сюда относятся рентгеноструктурный анализ и
электронная микроскопия, методы ЯМР и ЭПР, диффузное рассеяние рентг. лучей,
оптич. методы (исследование спектров поглощения, оптич. активности, люминесценции
и др.), микрокалориметрия, гидродинамич. методы, хроматография, электрофорез,
полярография и др. Изучение фотохим. и радиац--хим. изменений в П. б. служит
для исследования их структуры и для исследования механизма действия УФ- и ионизирующего
излучений на эти объекты. П. б. являются диэлектриками и полиэлектролитами,
поэтому важны измерения диэлектрич. поляризации и потерь в широком диапазоне
частот. Особый интерес представляет исследование кон-формац. превращений П.
б. в растворе, с этой целью используют спектрофотометрию в УФ-области и измерения
кругового дихроизма. В полипептидах при образовании из беспорядочного клубка
упорядоченной спиральной структуры в области длин волннм
наблюдается сильный гипохромный эффект (уменьшение поглощения), пригодный для
определения степени спи-ральности. Ароматические аминокислотные остатки имеют
полосы поглощения в областинм,
изменяющиеся при изменении окружения (неполярного на полярное), что позволяет
судить о расположении и контактах этих остатков в молекуле белка. Межплоскостные
взаимодействия в НК обусловливает гипохромный эффект в областинм.
Соответственно при разрушении двойной спирали (переходе спираль - клубок) наблюдается
увеличение поглощения на 40%. Прирост поглощения пропорционален доле нуклеотидов,
перешедших из упорядоченной спиральной структуры в неупорядоченный клубок. П.
б. обладают оптич. активностью, свойственной всем аминокислотам (кроме глицина)
и, соответственно, полипептидам и белкам. Наиб. информативны измерения кругового
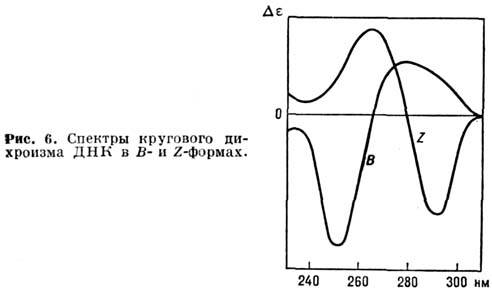
дихроизма, к-рый зависит от конформации полимера. На рис. 6 приведены кривые
кругового дихроизма для ДНК в b- и Z-формах.
Переходы спираль - клубок в П. б. Полипептидные
цепи, образующие в определ. условиях упорядоченные спиральные структуры, при
изменении внеш. условий переходят в состояние неупорядоченного клубка. Эти конформац.
переходы наиб. детально изучены на спн-тотич. гомогенных полипептидах. Переход
a-спираль - клубок носит кооперативный характер и характеризуется сравнительно
узким интервалом перехода. Коопе-ративность перехода обусловлена невыгодностью
освобождения из спиральной структуры (плавления) коротких участков, т. к. при
этом затрачивается значит. энергия на разрыв водородных связей, а выигрыш в
энтропии за счёт появления подвижности пептидных звеньев мал. При плавлении
длинных участков спирали возможна компенсация энергетич. затрат. Процесс денатурации
белков при изменении внеш. условий включает в себя и переход спираль - клубок,
но обычно процесс является многостадийным. Отд. стадии могут носить кооперативный
характер. Изучение промежуточных стадий и кинетики прямого и обратного процессов
(ренатурации) является источником сведений о самоорганизации высших структур
белковых глобул. Двойная спираль ДНК может разрушаться при изменении внеш. условий,
молекула при этом переходит в состояние одного или двух беспорядочных клубков
(при полном разделении нитей). Этот переход, также наз. переходом спираль -
клубок или внутримолекулярным плавлением, изучен экспериментально и теоретически
для b-формы ДНК. Переход спираль - клубок рассматривают на основе одномерной
Изинга модели.
В рамках модели объясняются все наблюдаемые на
опыте закономерности перехода в ДНК. Переход спираль - клубок в ДНК аналогичен
фазовому переходу 1-го рода, но не является истинным фазовым переходом, т. к.
молекулу можно рассматривать как одномерную систему. Интервал перехода (напр.,
интервал температур перехода) конечен. В этом интервале молекула разбивается на
чередующиеся спиральные и клубкообразные участки. Т. к. локальное или полное
разделение нитей двойной спирали ДНК происходит при мн. генетич. процессах в
клетке, причём в этом процессе участвуют др. молекулы, взаимодействующие с ДНК,
теория перехода спираль - клубок, включающая вопрос о влиянии др. молекул ("теорию
скрепок"), важна для понимания механизма функционирования ДНК.
ДНК в клетке обладает отрицат. сверхспирализацией,
т. е. двойная спираль в ней несколько раскручена (в кольцевых ДНК при этом двойная
спираль образует витки сверхспирали). В клетке есть система ферментов (топоизомераз),
изменяющих сверхспи-рализацию. Широко распространена лишь отрицат. сверхспирализация.
Сверхспиральная ДНК обладает повыш. энергией; топоизомеразы расходуют энергию
на создание сверхспирализации. Мерой сверхспирализа-ции является плотность сверхвитков
s (число сверхвитков, приходящееся на один виток двойной спирали). Величина
s отрицательна, ниже подразумевается её абс. значение. С ростом s
молекула ДНК становится более подвижной, реакционноспособной, увеличивается
вероятность нарушений структуры двойной спирали (локальных её раскрытий), в
отд. областях молекулы при достаточно большом значении s возникают альтернативные
(т. е. отличные от 5-формы) структуры - крестообразные структуры, Z- и Н- формы и др. Все эти структуры не образуются в линейной ДНК в
стандартных условиях. Энергия, необходимая для их образования, черпается из
энергии сверхспирализации. Для исследования альтернативных структур ДНК и определения
их энергетич. параметров используют эксперименты, анализируемые с помощью топологич.
теории. Топологич. ограничения, накладываемые кольцевым замкнутым строением,
приводят и к др. изменениям структуры и физ. свойств молекул ДНК. Исследование
влияния топологич. эффектов на строение и свойства ДНК и её биол. функции, на
регуляцию генетич. процессов является одной из задач молекулярной биофизики.
Литература по биологическим полимерам
Аккерман Ю., Биофизика, пер. с англ., М., 1964;
Физические методы исследования белков и нуклеиновых кислот, М., 1967;
Веденов А. А., Дыхне А. М., Франк-Каменецкий М. Д., Переход спираль - клубок в ДНК, "УФН", 1971, т. 105, в. 3, с. 479;
Блюменфельд Л. А., Проблемы биологической физики, 2 изд., М., 1977;
Шабарова 3. А., Богданов А. А., Химия нуклеиновых кислот и их компонентов, М., 1978;
Лазуркин Ю. С., Молекулярное плавление ДНК и эффект тонкой структуры кривых плавления, "Молекулярная биология", 1977, т. 11, в. 6, с. 1311;
Волькенштейн М. В., Биофизика, 2 изд., М., 1988,;
Франк-Каменецкий М. Д., Вологодский А. В., Топологические аспекты физики полимеров: теория и ее биофизические приложения, "УФН", 1981, т. 134, в. 4, с. 641;
Кантор Ч., Шиммел П., Биофизическая химия, пер. с англ., т. 1-3, М., 1984-85; "В мире науки", 1985, в. 12 (тематич. вып.);
Молекулярная биология клетки, пер. е англ., т. 1-5, М., 1986-87;
Вологодский А. В., Топология и физические свойства кольцевых ДНК, М., 1988.
Знаете ли Вы, в чем фокус эксперимента Майкельсона?
Эксперимент А. Майкельсона, Майкельсона - Морли - действительно является цирковым фокусом, загипнотизировавшим физиков на 120 лет.
Дело в том, что в его постановке и выводах произведена подмена, аналогичная подмене в школьной шуточной задачке на сообразительность, в которой спрашивается: - Cколько яблок на березе, если на одной ветке их 5, на другой ветке - 10 и так далее При этом внимание учеников намеренно отвлекается от того основополагающего факта, что на березе яблоки не растут, в принципе.
В эксперименте Майкельсона ставится вопрос о движении эфира относительно покоящегося в лабораторной системе интерферометра. Однако, если мы ищем эфир, как базовую материю, из которой состоит всё вещество интерферометра, лаборатории, да и Земли в целом, то, естественно, эфир тоже будет неподвижен, так как земное вещество есть всего навсего определенным образом структурированный эфир, и никак не может двигаться относительно самого себя.
Удивительно, что этот цирковой трюк овладел на 120 лет умами физиков на полном серьезе, хотя его прототипы есть в сказках-небылицах всех народов всех времен, включая барона Мюнхаузена, вытащившего себя за волосы из болота, и призванных показать детям возможные жульничества и тем защитить их во взрослой жизни. Подробнее читайте в FAQ по эфирной физике.
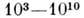 а.
е. м)., к ним приложимы все закономерности, установленные для др. природных
и синтетич. полимеров. Однако особенности хим. строения приводят к появлению
у П. б. уникальной пространств, структуры, необычных физ., хим. и биол. свойств.
По строению осн. цепи белки и НК однородны, подобно г о-мополимерам, у к-рых
все мономерные звенья цепи идентичны. Но в последовательности боковых групп
у П. б. закодирована генетич. информация организма, поэтому П. б. следует отнести
к гетеропо-лимерам с заданной нерегулярной последователь-ностью мономерных звеньев.
В структуре и свойствах П. б. отражены эти особенности их хим. строения. Пространств.
строение П. б. с определ. структурой всей макромолекулы наз. конформацией; от
конформации зависит взаимодействие П. б. с др. молекулами. Наиб. важные биол.
функции П. б. также определяются его конформацией и способностью изменять её при
разл. взаимодействиях. В большинстве случаев взаимодействия П. б. являются специфически-м
и, т. е. зависят от последовательности мономерных звеньев и локальной структуры
(см. также Биофизика).
а.
е. м)., к ним приложимы все закономерности, установленные для др. природных
и синтетич. полимеров. Однако особенности хим. строения приводят к появлению
у П. б. уникальной пространств, структуры, необычных физ., хим. и биол. свойств.
По строению осн. цепи белки и НК однородны, подобно г о-мополимерам, у к-рых
все мономерные звенья цепи идентичны. Но в последовательности боковых групп
у П. б. закодирована генетич. информация организма, поэтому П. б. следует отнести
к гетеропо-лимерам с заданной нерегулярной последователь-ностью мономерных звеньев.
В структуре и свойствах П. б. отражены эти особенности их хим. строения. Пространств.
строение П. б. с определ. структурой всей макромолекулы наз. конформацией; от
конформации зависит взаимодействие П. б. с др. молекулами. Наиб. важные биол.
функции П. б. также определяются его конформацией и способностью изменять её при
разл. взаимодействиях. В большинстве случаев взаимодействия П. б. являются специфически-м
и, т. е. зависят от последовательности мономерных звеньев и локальной структуры
(см. также Биофизика).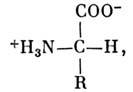
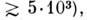 его индивидуальность определяется последовательностью
аминокислотных остатков. По своим функциям белки делятся на каталитические (ферменты,
биол. катализаторы хим. реакций), структурные, транспортные (гемоглобин), рецепторные,
ре-гуляторные (гормоны), защитные (антитела) и др. В зависимости от состава
выделяют простые белки- протеины, состоящие только из аминокислот, и сложные
белки -протеиды, в состав к-рых наряду с аминокислотами входят углеводы (гликопротеиды),
липида (липопротеиды), НК (нуклеопротеиды) и т.
д. По форме различают глобулярные белки, образующие плотные глобулы, и фибриллярные
белки, образующие длинные волокна или слои. Белки участвуют в важнейших генетич.
и регуляторных процессах. Нек-рые структурные белки могут образовывать агрегаты
в виде волокон, трубочек, оболочек. Иногда один и тот же белок выполняет неск.
функций.
его индивидуальность определяется последовательностью
аминокислотных остатков. По своим функциям белки делятся на каталитические (ферменты,
биол. катализаторы хим. реакций), структурные, транспортные (гемоглобин), рецепторные,
ре-гуляторные (гормоны), защитные (антитела) и др. В зависимости от состава
выделяют простые белки- протеины, состоящие только из аминокислот, и сложные
белки -протеиды, в состав к-рых наряду с аминокислотами входят углеводы (гликопротеиды),
липида (липопротеиды), НК (нуклеопротеиды) и т.
д. По форме различают глобулярные белки, образующие плотные глобулы, и фибриллярные
белки, образующие длинные волокна или слои. Белки участвуют в важнейших генетич.
и регуляторных процессах. Нек-рые структурные белки могут образовывать агрегаты
в виде волокон, трубочек, оболочек. Иногда один и тот же белок выполняет неск.
функций.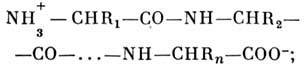
 спирали
и
спирали
и  структуры. a-Спираль
представляет собой правую спираль, у к-рой на один виток приходится 3,6 аминокислотных
остатка; шаг спирали 5,4
структуры. a-Спираль
представляет собой правую спираль, у к-рой на один виток приходится 3,6 аминокислотных
остатка; шаг спирали 5,4  ,
диаметр
,
диаметр (без боковых
групп).
(без боковых
групп).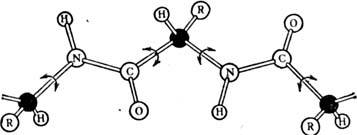
 спираль
представляет собой довольно жёсткую структуру. Не всем аминокислотным остаткам
энергетически выгодно образование a -
спирали. Знание соответствующих энергетич. параметров позволяет предсказывать
вероятность образования a-
спирали в том или ином участке белка. Существуют b-слои двух типов: параллельные
и антипарал-лельные. На рис. 2,б показана структура антипарал-лельного
спираль
представляет собой довольно жёсткую структуру. Не всем аминокислотным остаткам
энергетически выгодно образование a -
спирали. Знание соответствующих энергетич. параметров позволяет предсказывать
вероятность образования a-
спирали в том или ином участке белка. Существуют b-слои двух типов: параллельные
и антипарал-лельные. На рис. 2,б показана структура антипарал-лельного слоя.
Стабилизирующие
слоя.
Стабилизирующие слой
водородные связи между пептидными группами
направлены поперёк цепей, а сами цепи вытянуты и образуют складчатую структуру.
В белке встречаются также т. н.
слой
водородные связи между пептидными группами
направлены поперёк цепей, а сами цепи вытянуты и образуют складчатую структуру.
В белке встречаются также т. н.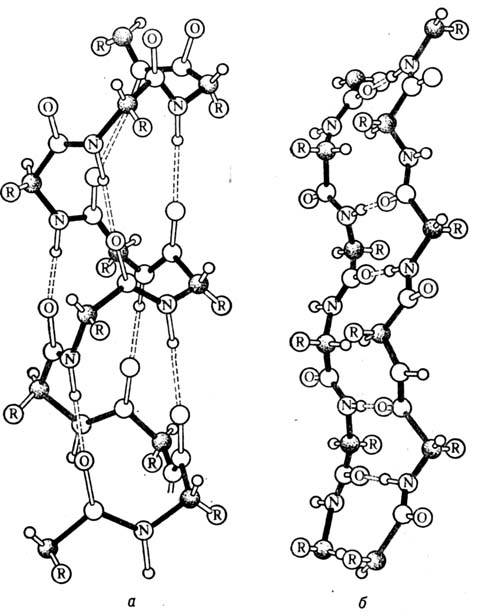
 при образовании
водородной связи. Возможны и др. типы спиралей. Все названные вторичные структуры
характерны для глобулярных белков. Фибриллярный белок, из к-рого строятся длинные
ориентиров. волокна, образует спирали иного вида. Вторичную (и третичную) структуру
белка исследуют с помощью
при образовании
водородной связи. Возможны и др. типы спиралей. Все названные вторичные структуры
характерны для глобулярных белков. Фибриллярный белок, из к-рого строятся длинные
ориентиров. волокна, образует спирали иного вида. Вторичную (и третичную) структуру
белка исследуют с помощью  спиралями.
Установлена закономерность в аминокислотной последовательности в этих
спиралями.
Установлена закономерность в аминокислотной последовательности в этих спиралях:
каждое 3-е или 4-е положение вдоль цепи занимают неполярные аминокислотные остатки.
При этом на боковой поверхности цилиндра, к-рым можно представить
спиралях:
каждое 3-е или 4-е положение вдоль цепи занимают неполярные аминокислотные остатки.
При этом на боковой поверхности цилиндра, к-рым можно представить 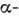 спираль,
образуется неполярная полоса, параллельная её оси. Именно эта гидрофобная полоса
обращена внутрь глобулы и контактирует с её гидрофобной частью.
спираль,
образуется неполярная полоса, параллельная её оси. Именно эта гидрофобная полоса
обращена внутрь глобулы и контактирует с её гидрофобной частью.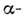 спиралей,
спиралей,
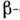 слоев и др.
элементов вторичной структуры. В этом случае можно говорить как о третичной
структуре таких доменов, так и о третичной структуре белков в целом, понимая
под этим взаимное расположение доменов в пространстве. Примером домена, содержащегося
во мн. белках, является блок из двух
слоев и др.
элементов вторичной структуры. В этом случае можно говорить как о третичной
структуре таких доменов, так и о третичной структуре белков в целом, понимая
под этим взаимное расположение доменов в пространстве. Примером домена, содержащегося
во мн. белках, является блок из двух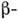 слоев,
соединённых между собой
слоев,
соединённых между собой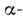 спиральным
сегментом. Доменная структура белков важна для их биол. функций. Вероятно также,
что домены - это элементарные белки, на основе к-рых в ходе эволюции возникает
разнообразие белковых структур.
спиральным
сегментом. Доменная структура белков важна для их биол. функций. Вероятно также,
что домены - это элементарные белки, на основе к-рых в ходе эволюции возникает
разнообразие белковых структур.
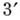 -и
атом С одного мономера соединяется фос-фодиэфирной связью с
-и
атом С одного мономера соединяется фос-фодиэфирной связью с 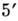 -м
атомом С следующего мономера. Каждая мономерная группа цепи ионизована p
несёт один отрицат. заряд. Размеры молекул РНК и ДНК изменяются в широких пределах.
Транспортные РНК (самые короткие молекулы РНК) состоят из 75-84 нуклеотидов;
длина гетерогенных ядерных РНК достигает
-м
атомом С следующего мономера. Каждая мономерная группа цепи ионизована p
несёт один отрицат. заряд. Размеры молекул РНК и ДНК изменяются в широких пределах.
Транспортные РНК (самые короткие молекулы РНК) состоят из 75-84 нуклеотидов;
длина гетерогенных ядерных РНК достигает 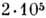 нуклеотидов. Короткие ДНК содержат обычно
неск. тысяч пар нуклеотидов, но существуют ДНК, к-рые содержат их
нуклеотидов. Короткие ДНК содержат обычно
неск. тысяч пар нуклеотидов, но существуют ДНК, к-рые содержат их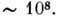
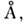 что оптимально
для межплоскостных взаимодействий, вносящих наиб. энергетич. вклад в стабильность
двойной спирали. В растворе на виток двойной спирали в
B-форме приходится 10,5 пары оснований. Диаметр двойной спирали равен примерно
22
что оптимально
для межплоскостных взаимодействий, вносящих наиб. энергетич. вклад в стабильность
двойной спирали. В растворе на виток двойной спирали в
B-форме приходится 10,5 пары оснований. Диаметр двойной спирали равен примерно
22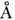 . В-форма
характерна для натриевой соли ДНК. При изменении внеш. условий (температуры, ионного
состава среды) параметры двойной спирали в В-форме изменяются, поэтому
следует говорить о 5-семействе структур. К этому семейству относится и литиевая
соль ДНК, т. н. С-форма, в к-рой на виток двойной спирали приходится 9,3 пары
оснований, плоскость оснований отклонена на
. В-форма
характерна для натриевой соли ДНК. При изменении внеш. условий (температуры, ионного
состава среды) параметры двойной спирали в В-форме изменяются, поэтому
следует говорить о 5-семействе структур. К этому семейству относится и литиевая
соль ДНК, т. н. С-форма, в к-рой на виток двойной спирали приходится 9,3 пары
оснований, плоскость оснований отклонена на 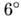 от
плоскости, перпендикулярной к оси спирали.
от
плоскости, перпендикулярной к оси спирали.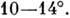 B-форма - осн. структура ДНК в живой клетке. ДНК может существовать и в др.
форме, в виде Z-спирали. Рентге-ноструктурный анализ позволил, как и в случае
белков, установить с высоким разрешением пространств.
структуры полинуклеотидов с разл. последовательностями нуклеотидов. Z-форма
ДНК, получившая своё назв. в связи с зигзагообразным строением сахарофосфатного
остова, представляет собой левую двойную спираль с периодом
B-форма - осн. структура ДНК в живой клетке. ДНК может существовать и в др.
форме, в виде Z-спирали. Рентге-ноструктурный анализ позволил, как и в случае
белков, установить с высоким разрешением пространств.
структуры полинуклеотидов с разл. последовательностями нуклеотидов. Z-форма
ДНК, получившая своё назв. в связи с зигзагообразным строением сахарофосфатного
остова, представляет собой левую двойную спираль с периодом 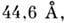 содержащую 12 пар оснований на виток и образованную антипараллельными полинуклеотидными
цепями, спаренными по правилам комплементарности. Повторяющимся звеном в ней
является не одна пара нуклеотидов, а две. Наиб. легко в Z-форму переходят регулярно
чередующиеся последовательности пурино-вых и пиримидиновых нуклеотидов. В физиологич.
условиях Z-форма в линейных ДНК не наблюдалась. Однако в кольцевых молекулах
ДНК может происходить переход отд. участков молекулы в Z-форму. На рис. 5 приведены
объёмные модели ДНК в В- и Z-формах.
содержащую 12 пар оснований на виток и образованную антипараллельными полинуклеотидными
цепями, спаренными по правилам комплементарности. Повторяющимся звеном в ней
является не одна пара нуклеотидов, а две. Наиб. легко в Z-форму переходят регулярно
чередующиеся последовательности пурино-вых и пиримидиновых нуклеотидов. В физиологич.
условиях Z-форма в линейных ДНК не наблюдалась. Однако в кольцевых молекулах
ДНК может происходить переход отд. участков молекулы в Z-форму. На рис. 5 приведены
объёмные модели ДНК в В- и Z-формах.
 нм
наблюдается сильный гипохромный эффект (уменьшение поглощения), пригодный для
определения степени спи-ральности. Ароматические аминокислотные остатки имеют
полосы поглощения в области
нм
наблюдается сильный гипохромный эффект (уменьшение поглощения), пригодный для
определения степени спи-ральности. Ароматические аминокислотные остатки имеют
полосы поглощения в области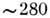 нм,
изменяющиеся при изменении окружения (неполярного на полярное), что позволяет
судить о расположении и контактах этих остатков в молекуле белка. Межплоскостные
взаимодействия в НК обусловливает гипохромный эффект в области
нм,
изменяющиеся при изменении окружения (неполярного на полярное), что позволяет
судить о расположении и контактах этих остатков в молекуле белка. Межплоскостные
взаимодействия в НК обусловливает гипохромный эффект в области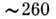 нм.
Соответственно при разрушении двойной спирали (переходе спираль - клубок) наблюдается
увеличение поглощения на 40%. Прирост поглощения пропорционален доле нуклеотидов,
перешедших из упорядоченной спиральной структуры в неупорядоченный клубок. П.
б. обладают оптич. активностью, свойственной всем аминокислотам (кроме глицина)
и, соответственно, полипептидам и белкам. Наиб. информативны измерения кругового
нм.
Соответственно при разрушении двойной спирали (переходе спираль - клубок) наблюдается
увеличение поглощения на 40%. Прирост поглощения пропорционален доле нуклеотидов,
перешедших из упорядоченной спиральной структуры в неупорядоченный клубок. П.
б. обладают оптич. активностью, свойственной всем аминокислотам (кроме глицина)
и, соответственно, полипептидам и белкам. Наиб. информативны измерения кругового