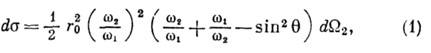
Клейна - Нишины формула - выражение для дифференц. сечения
ds рассеяния фотона на электроне (см. Комптона эффект ).В лаб. системе координат
где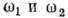 - частоты падающего и рассеянного фотона,
- частоты падающего и рассеянного фотона, 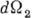 - элемент телесного угла для рассеянного фотона,
- элемент телесного угла для рассеянного фотона, 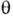 - угол рассеяния, параметр r0 = e2/mc2 =
2,81
- угол рассеяния, параметр r0 = e2/mc2 =
2,81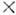
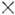 10-13 см - т. н. классический радиус электрона (е, т - заряд и масса электрона, с - скорость света).
10-13 см - т. н. классический радиус электрона (е, т - заряд и масса электрона, с - скорость света).
Частоты 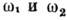 связаны соотношением Комптона:
связаны соотношением Комптона:
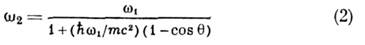
(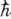 - постоянная Планка). Ф-лу (1) впервые получили О. Клейн и И. Нишяна (О. Klein,
I. Nishina) в 1929 в рамках теории, использующей формальный аппарат квантовой
механики И Дирака уравнение для описания релятивистского электрона. В
1930 эта ф-ла была заново выведена И. Е. Таммом.
- постоянная Планка). Ф-лу (1) впервые получили О. Клейн и И. Нишяна (О. Klein,
I. Nishina) в 1929 в рамках теории, использующей формальный аппарат квантовой
механики И Дирака уравнение для описания релятивистского электрона. В
1930 эта ф-ла была заново выведена И. Е. Таммом.
В пределе 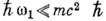 выпадает из ф-л (1) и (2), при этом (1) переходит в классич. ф-лу Томсона, описывающую
рассеяние света без изменения частоты (т. н. том-соновское рассеяние):
выпадает из ф-л (1) и (2), при этом (1) переходит в классич. ф-лу Томсона, описывающую
рассеяние света без изменения частоты (т. н. том-соновское рассеяние):
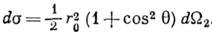
сти внутр. цилиндрич. канала
в молекуле бактериоро-допсина имеются гидрофильные аминокислоты. Внутри канала
- вода, связанная с гидрофильными аминокислотами водородными связями. В водном
канале молекулы бактериородопсина осуществляется первая стадия процесса фотосинтеза
в пурпурной мембране - превращение энергии света, поглощённого светочувствит.
группой бактериородопсина - ретиналем, в энергию переноса протона с внутренней
поверхности мембраны (из цитоплазмы)
во внешнюю среду.
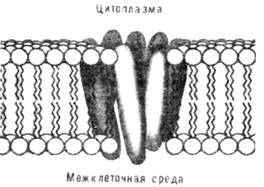
Рис. 2. Изображение структуры
белка бактериородопсина, встроенного в липидный слой.

Рис. 3. Регулярная укладка частиц на внутренней поверхности мембраны.
Фотосинтезиру ю щ а я система
зелёных растений (а также нек-рых бактерий и водорослей) образована белково-ли-пидными
мембранами, с к-рыми связаны молекулы пигмента - хлорофилла (в клетках растений
и бактерий встречаются разл. разновидности этого пигмента). Как и молекулы липидов,
хлорофилл имеет "полярную" головку (порфириновое кольцо, в состав
к-рого входит Mg2+ ) и "жирный хвост" (углеводородная
цепь). В фотосинтезирующей мембране гидрофобные хвосты хлорофилла погружены
в липидный слой, а полярные головки образуют монослой на поверхности мембраны.
Молекулы хлорофилла и мембранных белков образуют комплексы, к-рые упаковываются
в мембранах в регулярную двумерную решётку (рис. 3). В свою очередь, мембраны
уложены в слоистую структуру (аналогичную структуре лиотропных жидких кристаллов),
образуя т. н. граны. В процессе развития слоистая структура гран возникает из
скопления мембранных трубочек, аналогичного гексагональной фазе лиотрон-ного
жидкого кристалла (рис. 4). В мембранах гран расположены белковые комплексы,
осуществляющие последовательность хим. реакций, в результате к-рых энергия света,
поглощённого молекулами хлорофилла, преобразуется в химическую энергию АТФ (см.
Фотосинтез).
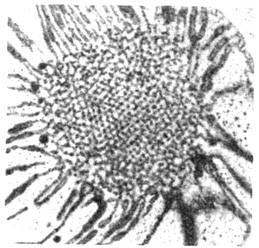
Рис. 4. Электронная микрофотография
слоистой структуры во время формирования фотосинтезирующей мембраны, 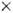 4000.
4000.
Специальной мембранной
структурой, улавливающей, как и хлоропласт, энергию света, являются фоторецепторы.
У позвоночных фоторецепторная система
(сетчатка глаза) образована слоем светочувствит. клеток (палочек и колбочек),
соединённых с нервными клетками (см. Зрение ).Палочки, необходимые для
зрения в условиях слабой освещённости, дают чёрно-белую картину, а менее чувствит.
колбочки обеспечивают цветовое зрение (имеется три типа колбочек, чувствительных
в разл. спектральных интервалах). Схема строения палочки изображена на рис.
5. Наружный членик окружён клеточной мембраной, внутри к-рой расположено ок.
500 параллельных дисков (их диаметр 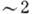 мкм, период структуры
мкм, период структуры 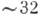 нм), каждый представляет собой двойную мембрану (уплощенный пузырек). Осн. белок
этой мембраны - родопсин (зрительный пурпур), имеющий светочувствит. группы
(хромофор) - ретиналь. Молекулы родопсина плотно упакованы в мембранах наружного
членика. В отличие от процессов, протекающих в хлоропластах, поглощение света
в мембранной структуре колбочек приводит не к синтезу АТФ, а к возбуждению нервного
импульса. Структура колбочек обеспечивает высокую чувствительность фоторецептор-ной
системы, способной реагировать на неск. квантов света.
нм), каждый представляет собой двойную мембрану (уплощенный пузырек). Осн. белок
этой мембраны - родопсин (зрительный пурпур), имеющий светочувствит. группы
(хромофор) - ретиналь. Молекулы родопсина плотно упакованы в мембранах наружного
членика. В отличие от процессов, протекающих в хлоропластах, поглощение света
в мембранной структуре колбочек приводит не к синтезу АТФ, а к возбуждению нервного
импульса. Структура колбочек обеспечивает высокую чувствительность фоторецептор-ной
системы, способной реагировать на неск. квантов света.
Внутренняя мембрана митохондрий.
Процесс синтеза АТФ, использующий хим. энергию, выделяющуюся при дыхании (т.
е. окислении органич. соединений) клеток, осуществляется в митохондриях, многочисл.
тельцах (размером 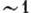 мкм), находящихся в цитоплазме эукариотич. клеток (т. е. клеток, имеющих ядро
и нек-рые др. органеллы). Митохондрия представляет собой мешок, образованный
двумя белково-липидны-ми мембранами (внеш. и внутр.), на внутр. мембране имеются
многочисл. складки - к р и с т ы (рис. 6), с к-рыми связаны ферменты, осуществляющие
процесс переноса электронов в реакции окисления. Белок цитохромоксидаза катализирует
реакцию окисления цитохрома (в результате на мембране создаётся градиент концентраций
прото нов, энергия к-рого используется для синтеза АТФ). Молекула цито-хромоксидазы
образована семью белковыми цепями, различными по размеру и составу аминокислот.
Погружённая в мембрану часть белка состоит из двух структурных доменов, соединённых
с третьим, выступающим в межмембранное пространство митохондрии, где фермент
взаимодействует с цитохромом. Отделённая от др. белков, цитохромоксидаза в окисленном
состоянии образует высокоупорядоч. структуры, к-рые разрушаются при восстановлении
этого фермента.
мкм), находящихся в цитоплазме эукариотич. клеток (т. е. клеток, имеющих ядро
и нек-рые др. органеллы). Митохондрия представляет собой мешок, образованный
двумя белково-липидны-ми мембранами (внеш. и внутр.), на внутр. мембране имеются
многочисл. складки - к р и с т ы (рис. 6), с к-рыми связаны ферменты, осуществляющие
процесс переноса электронов в реакции окисления. Белок цитохромоксидаза катализирует
реакцию окисления цитохрома (в результате на мембране создаётся градиент концентраций
прото нов, энергия к-рого используется для синтеза АТФ). Молекула цито-хромоксидазы
образована семью белковыми цепями, различными по размеру и составу аминокислот.
Погружённая в мембрану часть белка состоит из двух структурных доменов, соединённых
с третьим, выступающим в межмембранное пространство митохондрии, где фермент
взаимодействует с цитохромом. Отделённая от др. белков, цитохромоксидаза в окисленном
состоянии образует высокоупорядоч. структуры, к-рые разрушаются при восстановлении
этого фермента.
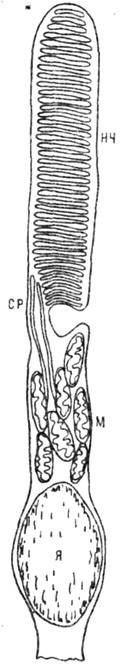
Рис. 5. Схема строения
палочки сетчатки позвоночных: НЧ- наружный
членик, СР - соединительная ресничка М- митохондрии,
Я - ядро.
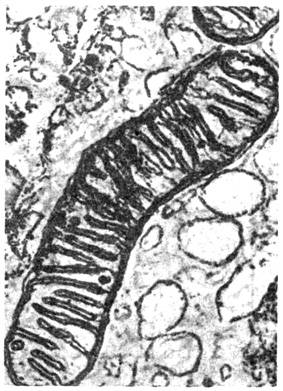
Рис. 6. Электронная микрофотография
митохондрии. Видны выросты на внутренней мембране - кристы.
Транспорт ионов и молекул.
Определенный хим. состав цитоплазмы каждой клетки (неравновесный по отношению
к внеш. среде) поддерживается регулированием транспорта разл. веществ через
мембраны, к-рый осуществляется через систему расположенных в мембранах каналов
(пассивный транспорт), молекул-переносчиков (облегченная диффузия) и насосов
(а к т и в н ы й транспорт), а также мета-болич. процессами и специфич. процессами
переноса крупных частиц сквозь мембраны (т. н. процессы эндо-и экзоцитоза).
Каналы образованы интегральными
белками, пронизывающими мембрану насквозь и имеющими форму тора; канал по отношению
к прохождению молекул может находиться в двух состояниях, открытом и закрытом.
Избирательность канала к разл. молекулам или ионам определяется его формой и
размерами, а также эл--статич. свойствами выстилающих его поверхность аминокислот.
На рис. 7 изображена трёхмерная структура мембранного белка (коннексона). Возможно
образование двумерной кристаллич. структуры коннексонов в области контакта мембран
двух клеток (рис. 8). Белок состоит из 6 одинаковых субъединиц, расположенных
таким образом, что в центре образуется канал (диаметром 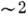 нм на внеш. стороне мембраны и сужающийся внутри). Часть молекулы этого белка
выступает из мембраны в межклеточное пространство, где она способна связываться
с коннексоном соседней клетки (образуя непрерывный канал, соединяющий внутр.
пространство двух клеток). Отд. субъединицы коннексона смыкаются, подобно лепесткам
диафрагмы фотоаппарата, при добавлении ионов Са2+ , закрывая просвет
канала и уменьшая его пропускную способность. Состояние нек-рых каналов зависит
также от величины разности потенциалов на мембране и наличия спец. хим. веществ
(медиаторов).
нм на внеш. стороне мембраны и сужающийся внутри). Часть молекулы этого белка
выступает из мембраны в межклеточное пространство, где она способна связываться
с коннексоном соседней клетки (образуя непрерывный канал, соединяющий внутр.
пространство двух клеток). Отд. субъединицы коннексона смыкаются, подобно лепесткам
диафрагмы фотоаппарата, при добавлении ионов Са2+ , закрывая просвет
канала и уменьшая его пропускную способность. Состояние нек-рых каналов зависит
также от величины разности потенциалов на мембране и наличия спец. хим. веществ
(медиаторов).
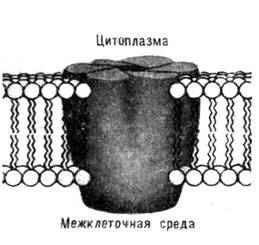
Рис. 7. Схема строения
клеточной мембраны со встроенными белками (коннексонами).

Рис. 8. Электронная микрофотография
кристаллической "бляшки", образованной белками (коннексонами) в
липидном слое.
Перенос веществ через каналы
происходит без затраты энергии и направлен в сторону уменьшения концентрации
молекул. Этим же свойством обладает т. н. процесс облегчённой диффузии, когда
молекула переносимого вещества связывается на одной стороне мембраны со спец.
молекулой-переносчиком в единый комплекс, к-рый легко проходит через мембрану,
а на другой её стороне распадается, отщепляя переносимую молекулу.
Системы активного транспорта
могут осуществлять перенос веществ против градиента концентрации (затрачивая
при этом энергию АТФ), обеспечивая в клетке оптимальную концентрацию ионов К+
и др. ионов, играющих важную роль в функционировании др. К. с., откачивая ионы
Na+, поддерживая пост. осмотич. давление
и т. д. Активный транспорт осуществляется специфич. мембранными белками за счёт
изменения третичной структуры при подведении к ним хим. энергии (молекулы АТФ)
или при поглощении кванта света. Самой распространённой системой активного транспорта
ионов является т. н. К+/Nа+ -активируемая АТФ-аза, откачивающая
ионы Na+ и одновременно обеспечивающая накопление ионов К+
в клетке, при этом одна молекула АТФ обеспечивает вывод трёх ионов Na+
из клетки и одноврем. ввод двух ионов К+. Большинство клеток содержит,
кроме того, насос др. типа, откачивающий ионы Na+ из клетки и обеспечивающий
движущую силу для активного переноса Сахаров и аминокислот в клетку.
Мн. клетки выкачивают из
цитоплазмы ионы Са2+ , расходуя при этом энергию АТФ. Активный транспорт
Са2+ осуществляется с помощью системы белковых субъединиц, включающей
регулируемый кальциевый канал, а также специфич. белок, изменяющий свою кон-формацию
(трёхмерную структуру) при связывании с ним иона Са2+ . Бактерии,
используя энергию метаболизма, создают в цитоплазме пониж. концентрацию протонов
при помощи спец. протонного насоса; при этом регулируется осмотич. давление
внутри клетки, а также поглощаются др. ионы, напр. К+ .
Аксоны нервных клеток,
имеющие развитую поверхность клеточной мембраны, окружены многослойной липидной
мембраной (миелиновой оболочкой), находящейся в жидкокристаллич. состоянии (рис.
9),
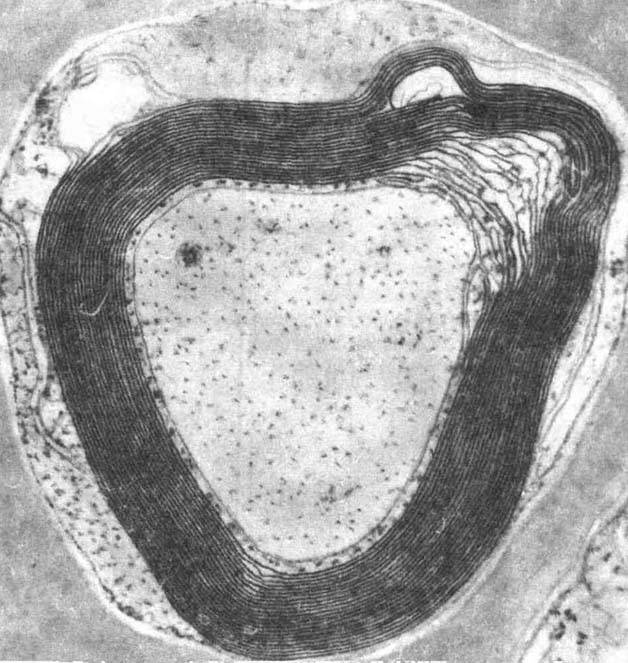
Рис. 9. Электронная микрофотография
поперечного среза нервного волокна, покрытого миелиновой оболочкой.
Такая оболочка уменьшает
пассивный выход натрия через плазматич. мембрану аксона, уменьшая тем самым
затраты на метаболизм нервной клетки и, в конечном итоге, увеличивая скорость
распространения нервного импульса.
Транспорт крупных частиц.
Синапс. Мн. клетки поглощают крупные частицы, вещество к-рых используется
в качестве строительного материала и источника энергии, при помощи процессов
эндоцитоза, включающих пиноцитоз (поглощение жидких капель) и фагоцитоз (поглощение
плотных частиц). При пиноцитозе клеточная мембрана, соприкоснувшись с частицей,
деформируется, образуя канал. Этот канал
заполняется жидкостью из внеш. среды, а затем отделяется и продвигается внутрь
цитоплазмы, где через нек-рое время мембрана растворяется и содержимое канала
высвобождается. При фагоцитозе частицу окружает вырост клеточной мембраны, к-рый
затем втягивается внутрь клетки. Образование и отделение каналов при зндоцитозе
требует затраты энергии клеткой.
"Узнавание"
внеклеточных частиц осуществляется с помощью системы рецепторов - белков, находящихся,
по крайней мере, часть времени вне клеточной мембраны, где они взаимодействуют
с внеш. раздражителями. Наряду со своей специфич. функцией, хемосорбцией, нек-рые
из этих белков играют роль молекул-переносчиков. Связав соответствующую молекулу,
белок-рецептор изменяет свою конформацию и стимулирует систему подвижности клетки.
Экзоцитоз (процесс, обратный
пиноцитозу) состоит в выделении во внеш. среду веществ, синтезированных в клетке
(напр., в мембранах т. н. комплекса Гольджи), в виде пузырьков жидкости. Подобный
процесс используется в спец. структуре (синапсе), осуществляющей передачу сигнала
от одной нервной клетки к другой с помощью выделения хим. вещества (медиатора).
Синапс образован мембранами двух соседних клеток, между к-рыми имеется зазор
(рис. 10). Вблизи первой мембраны внутри клетки расположено большое кол-во синаптич.
пузырьков, их поверхность образована двуслойной фосфолипидной мембраной, внутри
находится медиатор. Нервный импульс существенно изменяет ионный состав клетки:
ионы К+ замещаются на ионы Na+ , а также выделяется нек-рое
кол-во ионов Са2+ . Это приводит к встраиванию части пузырьков в
мембрану, при этом содержащийся в пузырьках медиатор выделяется в зазор между
клетками. Попадая затем на рецепторы второй мембраны, медиатор вызывает её возбуждение.
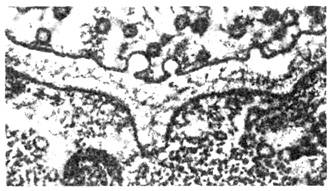
Ряс. 10. Электронная микрофотография
нервно-мышечного синапса лягушки в момент слияния синаптическо-го пузырька с
пре-синаптической мембраной и высвобождения медиатора.
2. Системы подвижности
клеток.
Передвижение клеток осуществляется
с помощью спец. систем, при этом расходуется часть энергии, получаемой клеткой
из процессов метаболизма, фотосинтеза и хемосинтеза. Бактерии движутся с помощью
нитевидных структур (жгутиков), соединённых с "мотором".
Бактериальный мотор состоит
(рис. 11) из системы колец (белковых частиц, имеющих форму диска) 4-5, к одному из к-рых прикреплён стержень, а остальные встроены в мембрану и стенку
бактериальной клетки 6-7, стенка служит для предохранения организма от
повреждений и представляет собой двумерную сеть, охватывающую клетку как мешок;
сеть образуется путём полимеризации углеводов и спец. белков клеточной стенки,
синтезированных в цитоплазме под действием ферментов, расположенных на внеш.
стороне клеточной мембраны. К стержню 3 прикреплён крюк 1, переходящий
в длинную нить (филамент) 2. Крюк и нить выступают из тела клетки и погружены
в окружающий раствор. Кольца, стержень, крюк и нить составляют бактериальный
жгутик. Обычно спираль жгутика является левовинтовой, и при нормальном прямолинейном
движении клетки эта спираль вращается против часовой стрелки.
Жгутиковые нити бактерий
обладают свойством самосборки. Диаметр нитей  13,5
нм, длина
13,5
нм, длина  неск.
мкм. Нити состоят из белка флагеллина, расположенного так, что филамент принимает
форму спирали, в к-рой на один виток приходится 8-10 белковых субъединиц. При
нагревании в растворе с физиологич. концентрацией соли нить диссоциирует; при
охлаждении такого раствора в присутствии затравочных структур (коротких фрагментов
нитей) происходит самосборка нитей. Образование крюка, стержня и колец бактериального
мотора также происходит путём самосборки из белковых субъединиц.
неск.
мкм. Нити состоят из белка флагеллина, расположенного так, что филамент принимает
форму спирали, в к-рой на один виток приходится 8-10 белковых субъединиц. При
нагревании в растворе с физиологич. концентрацией соли нить диссоциирует; при
охлаждении такого раствора в присутствии затравочных структур (коротких фрагментов
нитей) происходит самосборка нитей. Образование крюка, стержня и колец бактериального
мотора также происходит путём самосборки из белковых субъединиц.
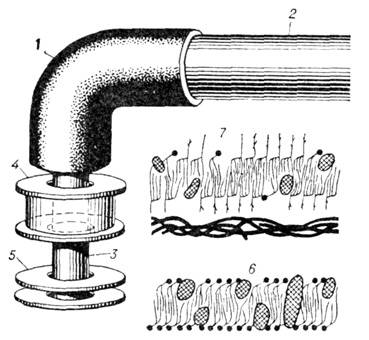
Рис. 11. Схема бактериального
мотора: 1 - крюк, 2 - нить (филамент), 3 - стержень, 4-5 - кольца, 6 - клеточная мембрана, 7 - клеточная оболочка.
У целого ряда бактерий
имеются только два кольца в бактериальном моторе: S-кольцо, прикреплённое
к стенке, и отстоящее от него на 3,5 нм М-кольцо, погружённое в клеточную мембрану
и скреплённое со стержнем мотора; кольца состоят из 15-16 секторов. В живой
бактериальной клетке часть энергии, получаемой ею при фотосинтезе и питании,
используется для приведения в действие протонного насоса, выкачивающего протоны
во внеш. среду и создающего в клетке пониж. концентрацию протонов. Предполагают,
что возникающий в результате этой разности концентраций поток протонов, проходящий
между S- и М-кольцами, приводит в действие бактериальный мотор; при этом
ротор мотора совершает один оборот относительно статора при прохождении примерно
300 протонов. Скорость вращения нити при работе мотора составляет обычно 10-
20 оборотов в секунду.
Микротубулярные структуры
эукариот. У эукарио-тич. клеток встречаются две системы подвижности: первая
строится из микротрубочек, осн. компонентом к-рых является белок тубулин, а
вторая - из микро-филаментов, содержащих гл. обр. актин. Микротрубочки связаны
с движением ресничек и жгутиков, миграцией пигментных гранул в хроматофорах
и т. п., микро-филаменты - с амебоидным движением, потоками цитоплазмы и т.
п. В системах подвижности кроме осн. белковых компонентов содержатся также др.
белки, к-рые непосредственно взаимодействуют с тубулином и актином при генерации
движения, выполняют регуля-торные функции, образуют соединения между микротрубочками,
микрофиламентами и др. структурами. Обе системы могут перекрываться, напр. разветвлённая
сеть микротрубочек внутри клеток может служить каркасом для прикрепления микрофиламентов
(рис. 12).
Микротрубочки представляют
собой длинные полые цилиндры, наружный диаметр к-рых  24
нм, внутренний
24
нм, внутренний  15
нм, длина
15
нм, длина  неск.
мкм (за исключением нек-рых клеток, напр. нейронов, достигающих длины в неск.
см). Микротрубочки осуществляют генерацию движения клеток, участвуют в поддержании
формы клетки, во внутриклеточном
транспорте веществ, в секреции клеточных продуктов (процессах экзоцитоза), в
движении хромосом клетки. Микротрубочки могут быть рассеяны в цитоплазме или
собраны в организов. структуры.
неск.
мкм (за исключением нек-рых клеток, напр. нейронов, достигающих длины в неск.
см). Микротрубочки осуществляют генерацию движения клеток, участвуют в поддержании
формы клетки, во внутриклеточном
транспорте веществ, в секреции клеточных продуктов (процессах экзоцитоза), в
движении хромосом клетки. Микротрубочки могут быть рассеяны в цитоплазме или
собраны в организов. структуры.
Реснички и жгутики эукариот
устроены след. образом: центр. пару микротрубочек окружает кольцо из микротрубочек,
обычно парных. Микротрубочки связаны между собой поперечными мостиками; вся
конструкция наз. аксонемой (рис. 13), Стенка каждой из центр. микротрубочек
образована 13 продольными рядами (протофибриллами) белковых субъединиц. В каждом
из девяти дублетов микротрубочек, окружающих центр, пару, одна (А)имеет
полностью замкнутую стенку, состоящую из 13 протофибрилл, а вторая (Б)имеет
серповидную форму (состоит из 10 протофибрилл) и примыкает к первой. Каждая
микротрубочка А соединена с микротрубочкой Б соседнего дублета.
От обращённой внутрь поверхности микротрубочки А отходит пo направлению
к центру белковый выступ - "радиальная спица", кроме того, каждая
микротрубочка А имеет боковые "ручки" длиной ~14 нм. Все
эти выступы расположены с определ. периодичностью и играют важную роль в генерации
движения. Спец. структура (базальное тельце), из к-рой исходит жгутик, находится
в цитоплазме клетки и действует как организатор ассоциации микротрубочек. Самосборка
микротрубочек имеет неск. стадий: сборка колец и фрагментов спиралей из тубулина,
формирование ленточных структур, свёртывание листовых структур в микротрубочки.
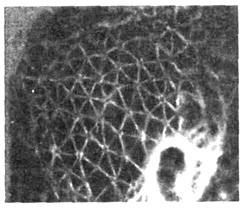
Рис. 12. Изображение трёхмерной
сети актиновых нитей в клетке, полученное с помощью флуоресцирующих антител,
специфично связывающихся с белком тропомио-зином.
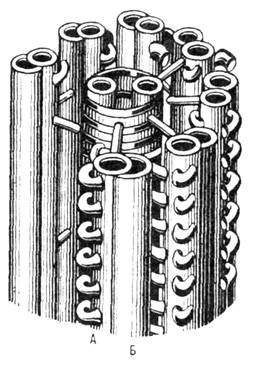
Рис. 13. Схематическое
изображение аксонемы жгутика.
Микротрубочки могут генерировать
движение с помощью двух разл. механизмов: за счёт активного скольжения (подобного
аналогичному процессу в мышечном волокне; см. ниже) или же путём изменения своей
длины вследствие полимеризации или деполимеризации микротрубочек. К последнему
типу относится движение хромосом.
Микрофиламенты эукариотич.
клеток представляют собой длинные нитевидные структуры толщиной 5-7 нм, находящиеся
в цитоплазме, и состоят гл. обр. из актина. Обычно из микрофиламентов образуются
подвижные пучки или тонкие сетчатые структуры, форма и локализация к-рых в цитоплазме
зависят от жизненного цикла клетки, её движения и др., а в нек-рых структурах,
напр. в мышечных волокнах,- упорядоченные и стабильные структуры. Когда клетки
находятся в состоянии покоя и прикреплены к стенкам сосуда, в к-ром живут, в
цитоплазме имеются длинные пучки микрофиламентов, располагающиеся под мембраной
(стрессовые волокна).


Рис. 14. Электронная микрофотография
продольного (сверху) и поперечного (внизу) срезов скелетной мышцы.
Нек-рые жгутиковые передвигаются с помощью спец. структуры, т. н. сократимого аксостиля, способного совершать волнообразные движения. Сократимый аксостиль - ленточная структура, проходящая от одного конца клетки до другого через всю цитоплазму. Каждый лист аксостиля построен из множества микротрубочек, число листов варьирует от 2-3 до 20-30, листы в аксостиле находятся на расстоянии ~30 нм друг от друга, а расстояние между соседними микротрубочками одного листа ~40 нм. Белковые мостики, расположенные в определ. порядке вдоль микротрубочек, соединяют их между собой в пределах каждого листа. Такая структура обладает подвижностью, т. к. построенные из микротрубочек листы могут скользить относительно друг друга. Мышечное волокно. Мышца представляет собой высокоупорядоченную структуру, состоящую из мышечных волокон (миофибрилл), к-рые образованы тонкими и толстыми нитями диаметром 3 и 10 нм соответственно, упакованными в гексагональную решётку (рис. 14). Тонкие нити образованы белком актином, а толстые - миозином. Одним своим концом актиновые нити прикреплены к т. н. Z-пластинкам, участок мышцы между двумя Z-пластинками наз. саркомером. Микроскопически структура миофибриллы напоминает гексагональную фазу лиотропного жидкого кристалла, в к-рой роль цилиндрич. мицелл играют актиновые и миозиновые нити. Период кристаллич. структуры в поперечном срезе мышцы определяется дальнодействую-щими ван-дер-ваальсовыми и эл--статич. силами; при увеличении концентрации ионов в пространстве между нитями (при пост, длине саркомера) период гексагональной решётки уменьшается. В естеств. условиях сокращение мышцы инициируется нервным импульсом, в ответ на к-рый нити актина и миозина скользят относительно друг друга, при этом их перекрытие увеличивается, а длина саркомера уменьшается. Активное скольжение нитей в мышечном волокне осуществляется с помощью т. н. головки молекулы миозина (рис. 15), к-рая может находиться в двух конформац. состояниях. Связав молекулу АТФ, миозиновая головка переходит в активированное состояние и прикрепляется к одной из актиновых субъединиц ближайшего микрофиламента, что, в свою очередь, вызывает гидролиз АТФ. За счёт выделившейся при этом энергии головка переходит в новое конформац. состояние и немного перемещает актиновый микрофиламент, к к-рому головка прикреплена. Связывание комплекса миозин-АТФ с актином возможно только в присутствии ионов Са2+ , к-рые высвобождаются при деполяризации мембраны, вызванной приходом нервного импульса.
Сборка актиновых и миозиновых
нитей может происходить из белковых субъединиц и вне живой клетки. В растворе
актин и миозин могут соединяться в комплекс - актомиозин, образующий гель. При
добавлении в систему АТФ этот гель способен сокращаться и совершать работу аналогично
живой мышце. По-видимому, этот процесс важен при движении микроорганизмов: периодическое
набухание и осушение актомиозинового геля вызывает движение цитоплазмы клетки
и её перемещение как целого (по подложке).
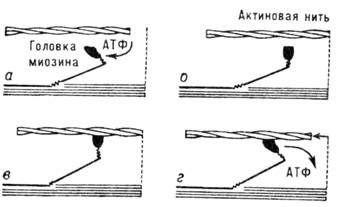
Рис. 15. Схематическое
изображение одного цикла взаимного проскальзывания актиновых и мио-зиновых нитей.
Присоединение молекулы АТФ к головке миозина (а) вызывает изменение её конформации
(б) и присоединение к актиновой нити (в). Гидролиз АТФ и её отсоединение от
головки приводят к
смещению актиновой нити (г) и восстановлению первоначальной конформации
головки миозина.
Митотическое веретено
- спец. К. с., участвующая в процессе деления (митоза) клетки. Веретено возникает
на определ. стадии клеточного цикла (рис. 16) и состоит из пучков волокон, образованных
микротрубочками и микрофиламентами. Волокна соединены с двумя тельцами - центриолями
(аналогичными базальным тельцам жгутиков микроорганизмов). X р о-м о с о м ы,
в к-рых хранится два экземпляра генетич. информации клетки (в двух сестринских
хроматидах), прикрепляются к нитям веретена, а затем сестринские хроматиды расходятся
к противоположным полюсам. На последней стадии в области соединения дочерних
клеток возникает кольцо, образованное нитями актина и миозина, обеспечивающее
разделение клеток.
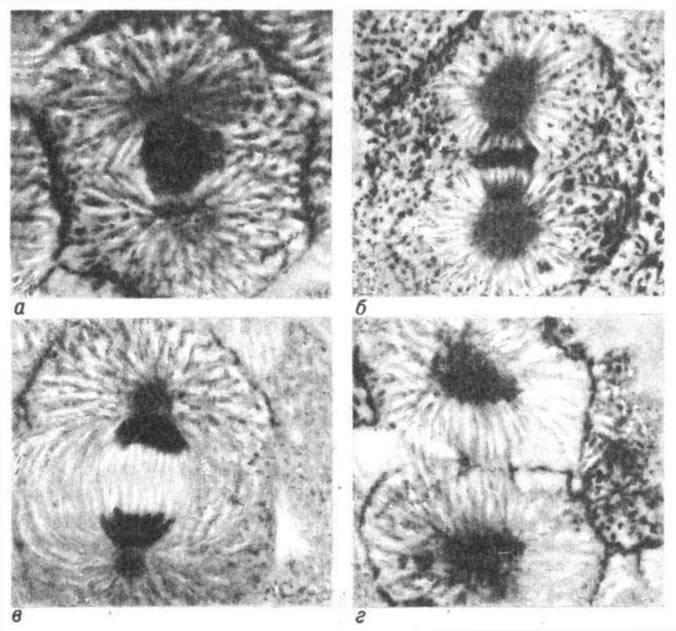
Рис. 16. Стадии
деления зародыша сига: а - профаза (видно начало
образования веретена); б - метафаза; в - анафаза; г - телефаза (кольцо разделяет клетку на две новые дочерние клетки).
3. Структура хромосом.
Вирусы.
Плотноупакованные хромосомы
представляют собой К. с., образованную белками (в т. ч. пятью белками, ваз.
гистонами, содержащими от 50 до 200 аминокисгидрофобных связей между субъединицами
приводит к образованию спирали, в центре к-рой находится цилиндрич. полость
диаметром ~2 ни. Структура белковых субъединиц тако- ва, что при их сборке образуется
спиральный желобок, закрученный вокруг длинной оси стержня. В этом желобке укладывается
одна одноцепочечная молекула рибонуклеиновой кислоты (РНК, в к-рой хранится
генетич. информация вируса) длиной ~6400 нуклеотидов (рис. 19). Частица BTiM
имеет длину 300 нм, диаметр ~18 нм.

Рис. 17. Электронная микрофотография кристаллической структуры, образованной ядрами нуклеосом, сверхспирали которых упакованы в гексагональную решётку.
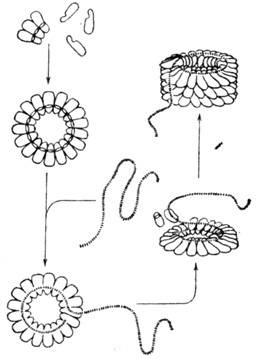
Рис. 18. Схематическое
изображение структуры ВТМ.
Рис. 19. Схема сборки частицы
ВТМ из изолированных субъединиц белка оболочки и молекулы РНК вируса.
Вещество вирусов синтезируется
с помощью мета-болич. аппарата клетки-хозяина; частицы вируса собираются в цитоплазме
клетки, иногда образуя упорядоченные структуры (рис. 20; см. Биологический
кристалл).
Хранение генетической информации
вирусов (не имеющих собств. метаболич. аппарата для воспроизведения своей структуры,
а использующих для этих целей структуры клеток, к-рые данный вирус инфицирует)
обеспечивается определ. структурой оболочки этих организмов. На рис. 18 приведено
схематич. изображение т. н. вируса табачной мозаики (ВТМ). Частица ВТМ состоит
из 2130 одинаковых молекул белка (длиной 158 аминокислот). Последовательная
агрегация этих удлинённых белковых молекул с помощью
Синтез белков в клетке
(в процессе к-рого используется генетич. информация, записанная в виде последовательности
нуклеотидных пар в ДНК) осуществляется в спец. комплексах - рибосомах, имеющих
размер ~25 нм. Рибосома состоит из двух субъединиц (рис. 21), к-рые образованы
молекулами РНК (меньшая из двух субъединиц содержит одну молекулу РНК, а большая
- две), и белков (субъединицы содержат по нескольку десятков белковых молекул).

Рис. 20. Электронная микрофотография
частиц вируса полиомиелита, плотно упакованных в цитоплазме клетки в кристаллическую
структуру,  200
000.
200
000.
Рис. 21. Схематическое изображение рибосомы.

А. А. Веденов, Е. Б. Левченко
Понятие же "физического вакуума" в релятивистской квантовой теории поля подразумевает, что во-первых, он не имеет физической природы, в нем лишь виртуальные частицы у которых нет физической системы отсчета, это "фантомы", во-вторых, "физический вакуум" - это наинизшее состояние поля, "нуль-точка", что противоречит реальным фактам, так как, на самом деле, вся энергия материи содержится в эфире и нет иной энергии и иного носителя полей и вещества кроме самого эфира.
В отличие от лукавого понятия "физический вакуум", как бы совместимого с релятивизмом, понятие "эфир" подразумевает наличие базового уровня всей физической материи, имеющего как собственную систему отсчета (обнаруживаемую экспериментально, например, через фоновое космичекое излучение, - тепловое излучение самого эфира), так и являющимся носителем 100% энергии вселенной, а не "нуль-точкой" или "остаточными", "нулевыми колебаниями пространства". Подробнее читайте в FAQ по эфирной физике.
|
|
