Г.Ю.Ризниченко.
Лекции
по математическим моделям в биологии
ЛЕКЦИЯ 7
МУЛЬТИСТАЦИОНАРНЫЕ СИСТЕМЫ
Триггер. Примеры систем с двумя
устойчивыми стационарными состояниями. Конкуренция. Силовое и параметрическое
переключение триггера. Эволюция. Отбор одного из двух и нескольких равноправных
видов. Генетический триггер Жакоба и Моно.
Важная особенность биологических систем – переключение из одного режима функционирования в другой. Приведем простые примеры переключения процессов в живых системах:
·
Сон и бодрствование – это разные
типы метаболизма. Переключение происходит периодически и синхронизируется
геофизическим ритмом.
·
У большинства насекомых один и тот же организм может
существовать в виде гусеницы, куколки, бабочки. Переключение происходит
последовательно в соответствии с генетической программой.
·
Дифференцировка тканей –
клетки получаются путем деления из одного типа клеток, но впоследствии каждая выполняет
свои функции.
На фазовой плоскости триггерной системе в простейшем
случае соответствует два или несколько устойчивых стационарных решений,
разделенных сепаратрисами. Напомним,
что все особые точки (устойчивые и седло) лежат на пересечении главных изоклин – изоклин вертикальных и горизонтальных касательных (см.
Лекция 4).
На рис. 7.1 представлен относительно
простой фазовый портрет триггерной системы, описывающей явление конкуренции
двух одинаковых видов:
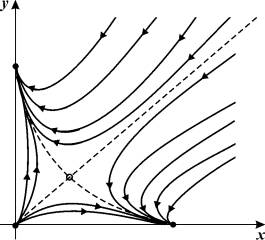
Рис. 7.1. Фазовый портрет триггерной
системы, описывающей явление конкуренции между двумя одинаковыми видами.
Соответствующая система уравнений имеет вид:
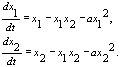
Такая
система имеет четыре стационарных решения:
1. x1=x2=0 – неустойчивый
узел;
2.
![]()
3.
![]()
4.
![]()
Бистабильная система может иметь гораздо более сложную
структуру фазового портрета. Пример такой системы ‑ движение шарика в
ложбине с двумя лунками в присутствии трения (Д.С. Чернавский).
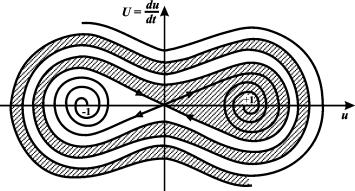
Рис. 7.2. Фазовый портрет системы 7.2 (шарик в ложбине с
двумя лунками). Темным обозначена область притяжения стационарного состояния
(+1)
Система описывается уравнениями:
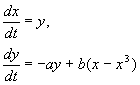
В
такой системе три стационарных состояния. Состояние x=y=0 – седло. Два
других стационарных состояния – устойчивые фокусы.
Вблизи этих стационарных состояний траектории представляют собой закручивающиеся
спирали. Вдали от стационарных состояний области притяжения имеют слоистую
структуру. Толщина слоев уменьшается при уменьшении параметра a.
Как видно из приведенных выше примеров, в триггерных
системах и поведение во времени и стационарное решение зависит не только от
параметров, но и от начальных условий.
Способы переключения триггера
Слово триггер означает
переключатель. Встает вопрос, как можно
переключить триггер из одного в другое стационарное состояние?
Рассмотрим фазовый портрет системы, обладающей двумя
устойчивыми стационарными состояниями (рис. 7.3). Здесь a,c – устойчивые стационарные состояния, b – седло.
Рис. 7.3. Триггерная система. Жирными линиями показаны
главные изоклины. Пунктирной линией обозначена сепаратриса, отделяющая
области влияния двух устойчивых стационарных состояний a и с
Двойная стрелка показывает процесс силового переключения триггера.
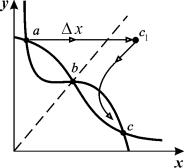
Если начальное положение изображающей точки расположено
левее сепаратрисы седла (пунктирная линия),
система находится в области притяжения особой точки a
и со временем стремится к этому устойчивому стационарному состоянию. Из точек,
лежащих правее сепаратрисы, система будет двигаться к особой точке c. Рассмотрим возможные способы переключения системы из
режима a в режим c.
1. Силовое переключение. Можно изменить значения концентраций (например, добавить
определенное количество вещества x1, так что система «перепрыгнет»
через сепаратрису, например в некоторую точку c1, которая находится по правую сторону сепаратрисы в
области влияния устойчивого стационарного состояния с, к которому система перейдет сама с течением времени. На фазовом
портрете рис. 7.3 силовое (специфическое) переключение показано двойной
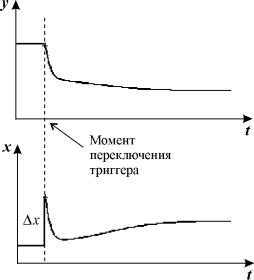
стрелкой. Кинетика переменных при таком переключении показана на рис. 7.4.
Рис. 7.4. Поведение переменных во времени при силовом переключении
после добавления в систему вещества x в количестве, достаточном для переключения системы из режима a в режим c
(смотри рис. 7.3).
2. Параметрическое переключение.
Другой
– неспецифический способ переключения показан на рис. 7.5, 7.6.
При таком способе переключения непосредственному
воздействию подвергаются не переменные, а параметры системы. Это может быть
достигнуто разными способами, например, изменением скорости поступления
субстрата, температуры, рН.
Сущность такого способа переключения состоит в
использовании зависимости фазового портрета системы от некоторого управляющего
параметра. Пусть с изменением этого параметра фазовый портрет претерпевает
последовательность превращений, показанных на рис. 7.5 (а – г).
На стадии (в) устойчивый узел (а) и
седло (b) сливаются в одну полуустойчивую точку седло‑узел.
На стадии (с) в системе остается лишь
одно устойчивое стационарное состояние, к которому и сходятся все фазовые
траектории.
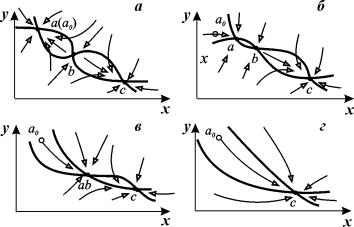
Рис. 7.5. Параметрическое переключение
триггера. Последовательные стадии трансформации фазового портрета. Стрелками
обозначено направление фазовых траекторий.
Рис. 7.6. Кинетика изменения переменных в процессе параметрического переключения
триггера.

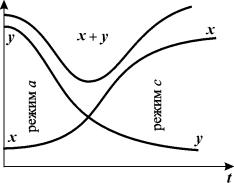
Тогда система, находившаяся в начале процесса
переключения в стационарном режиме а,
в результате параметрического переключения окажется в области притяжения
единственного устойчивого стационарного режима с, куда с течением времени и перейдет (рис. 7.6).
Параметрический способ переключения реализуется при изменении любой генетической программы, он может также иметь место при изменении внешних условий, приводящих к изменению управляющего параметра системы.
Модели отбора
Как мы видели выше, в триггерной системе изображающая
точка “выбирает” (в зависимости от параметров и начальных условий) стационарный
режим функционирования. Триггерные модели могут быть использованы при описании
процесса отбора, и потому применимы к процессам эволюции.
Геохронологическая таблица
|
1000 тыс. лет |
КАЙНОЗОЙ |
Эволюция человека |
|
|
МЕЗОЗОЙ |
Появление млекопитающих |
|
|
ПАЛЕОЗОЙ |
Первые многоклеточные |
|
2000 – 3000 тыс.
лет |
ПРОТЕОЗОЙ |
Биологическая эволюция |
|
4000 тыс. лет |
АРХЕЙ |
Микроископаемые |
|
5000 тыс. лет |
|
Образование земли |
|
6000 тыс. лет |
|
Возникновение солнечной
системы |
Изучая приведенную выше геохронологическую таблицу, можно
выделить два класса процессов эволюции:
1) Системы: где новые элементы не появляются, а старые не
исчезают – происходит их перераспределение в пространстве и во времени.
2) Возможен самопроизвольный отбор немногих элементов (и их размножение) из очень большого числа различных уже существующих или тех, которые могут возникнуть.
К первому типу относятся процессы
эволюции галактик, упорядоченных вихрей в гидродинамике, автоколебаний и
автоволн в активных средах. Сюда же относятся процессы образования негомогенных
стационарных распределений вещества в пространстве – диссипативных структур.
Более подробно эти процессы будут рассмотрены во второй части лекций.
Ко второму типу относится образование
изотопов химических элементов, макромолекул в химической эволюции и видов в
биологической эволюции, а также образование человеческих языков. Все эти
процессы идут в результате размножения и конкурентного отбора.
Структурирование в пространстве (тип 1)
обычно предшествует конкурентному отбору (тип 2). До образования изотопов
химических элементов (водород и гелий) должны были возникнуть "сгустки материи" ‑ зародыши
галактик и звезд. До возникновения макромолекул должна была образоваться
планетная структура и атмосфера Земли, доступная для солнечных лучей.
Человеческие языки возникают в замкнутых коллективах и проч.
Наоборот, процессы отбора ведут к возможности образования
новых, более сложных структур. На базе разнообразия макромолекул идет становление
живого организма. Биогеоценозы формируются из разных видов живых существ и т.д.
Главный вопрос эволюции: «как появилось свойство авторепродукции?» включает в себя несколько
вопросов. Вот основные из них:
·
Как возникли комплексы
белка и полинуклеотидов?
·
Как образовался единый
генетический код? (т.е. соответствие между последовательностями нуклеотидов в
ДНК и аминокислот в белках?)
·
Почему именно эти три
нуклеотида кодируют данную аминокислоту (кодон).
Действительно, существующий генетический код не связан с физико-химическими свойствами аминокислот и кодонов. Число равноправных кодов ‑ 20! а реализован только один. Вероятность случайного возникновения именно существующего кода крайне мала.
На вопрос «Как же произошел отбор»? возможно несколько ответов:
Кастлер: начальный код возник случайно, другие комбинации не успели возникнуть.
Эйген: возникло несколько разных кодов, но отобрались
наилучшие.
Чернавский: произошел отбор одного из равноправных.
Модель образования единого кода.
Можно выделить четыре
стадии эволюции формирования единого генетического кода.
1. Образование первичного бульона.
2. Образование белково-нуклеотидных комплексов,
способных к авторепродукции.
3. Образование единого кода в результате отбора.
4. Образование разных видов на основе единого
кода.
Рассмотрим 3-й этап. Мы уже говорили, что
существует три возможных механизма:
а)
Один объект возникает раньше других и развивается так быстро, что другие не
успевают возникнуть.
б) В результате конкуренции между объектами с различными
свойствами выжили и отобрались наилучшие, обеспечив наибольшую скорость
репликации.
в) В
результате антагонистического взаимодействия между равноправными объектами (с
одинаковой скоростью репликации), но разными последовательностями нуклеотидов,
выживает один вид объектов.
Действие каждого из этих механизмов может привести к
возникновению совокупности полностью одинаковых объектов, в которой одной
последовательности нуклеотидов соответствует одна последовательность
аминокислот ‑ однозначный код.
Отбор одного из
равноправных
Общая модель такого отбора имеет вид:
![]()
Здесь a - эффективный коэффициент репродукции, g - вероятность гибели в результате встречи.
Пусть N= 2, X1 = x, X2 = y. Система
уравнений имеет вид:
![]()
Стационарные решения находятся из алгебраических уравнений, полученных приравниванием правых частей нулю.
![]()
Система имеет два стационарных решения:

Для второго – нетривиального симметричного стационарного состояния характеристический определитель системы имеет вид:

Характеристическое
уравнение:
![]()
или
![]()
Выражения для характеристических чисел находятся из уравнения:
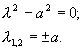
Корни положительны и разных знаков. Это означает, что
симметричное стационарное состояние представляет собой седло.
Аналогичный анализ показывает, что нулевая особая точка представляет собой неустойчивый узел.
Изоклины горизонтальных касательных: y=0 – ось абсцисс и вертикальная прямая x=a/g; изоклины вертикальных касательных: x=0 – ось ординат – и горизонтальная
прямая y=a/g.
Все траектории уходят на бесконечность, так как самоограничение роста популяции в данной модели не учитывается.
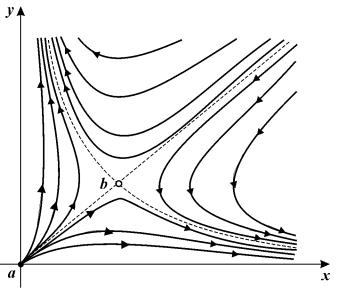
Рис. 7.7.
Фазовый портрет системы (7.4), описывающей отбор одного из двух
равноправных видов в отсутствие ограничений роста. а (начало координат) – неустойчивый узел, b – седло
Биологический
смысл модели.
Модель (7.4) демонстрирует принципиальную возможность отбора в системе равноправных видов, где симметричное состояние сосуществования является неустойчивым. Вот один из примеров такой системы.
Известно, что сахара и аминокислоты
являются оптически активными соединениями, причем сахара – левовращающие
плоскость поляризации света, аминокислоты – правовращающие. Противоположные
изомеры не только не встречаются в живых организмах и не усваиваются ими, но являются
ядами. В этом заключается одна из сложностей искусственного синтеза.
Ясно, что «зеркальные» организмы не лучше
и не хуже. В неживой природе распространены рацемические смеси, содержащие
равное количество зеркальных изомеров. То же – при небиологическим синтезе.
По-видимому, и первичный бульон был рацемической смесью.
Рассмотренная модель описывает выживание одних и
уничтожение других. Условие, которое обеспечивает при этом отбор одного вида,
заключается в том, что при встрече они взаимно отравляются и гибнут. Причина отбора здесь – не преимущество
одного из видов, а их взаимный антагонизм.
Однако модель (7.4) не может
описывать реальную систему, так как описывает неограниченный рост биомассы с
течением времени. Этот недостаток может быть исправлен несколькими способами.
Один их них – введение самоограничения численности вида в виде ферхюльстовских
членов. Тогда мы придем к модели (7.1). Другой способ – ввести в модель
переменную, описывающую поступающий в систему с определенной скоростью
питательный ресурс, общий для обоих видов.
Учтем ограниченность питательных ресурсов. Пусть S -лимитирующий субстрат (световая энергия, минеральное
питание и т.п.). Сам субстрат не является оптически активным, но преобразуется
в оптически активные продукты.
Выразим скорость роста каждой популяции a через S в соответствии с формулой Моно (7.9.). График этой
функции приведет на рис. 6.3.
![]()
Пусть n ‑ интенсивность притока субстрата. Расход
субстрата пропорционален поглощению его организмами, т.е. сумме их
концентраций. Уравнение для скорости изменения концентрации субстрата во
времени имеет вид:
![]()
Здесь
а > 1 - экономический
коэффициент - указывает, сколько субстрата идет на образование единицы биомассы.
Уравнения для концентраций объектов типа x и y:
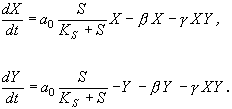
Введем безразмерные переменные:
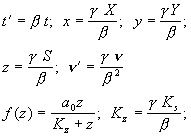
Система в безразмерном виде:
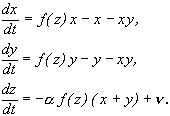
Пусть процессы поглощения субстрата существенно более
быстрые, чем процессы репродукции. В этом случае может быть использован метод
квазистационарных концентраций (лекция 6), и дифференциальное уравнение для быстрой
переменной z (S) – концентрации субстрата – заменено алгебраическим.
Тогда субстрат на интересующих нас временах достигнет квазистационарной
концентрации: dz/dt=0.
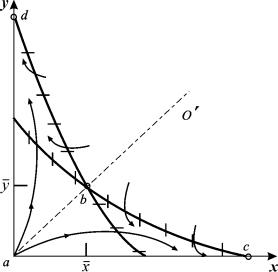
Рис. 7.8. Фазовый портрет системы 7.15,
описывающей отбор одного из двух равноправных видов когда субстрат поступает в
систему с постоянной скоростью. а (начало
координат) – неустойчивый узел, b – седло, c, d –устойчивые узлы.
Отсюда
![]()
В итоге получается система двух безразмерных уравнений
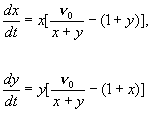
Построим фазовый портрет системы. (Рис.7.8) Изоклины вертикальных касательных:
x=0 (ось ординат) и
кривая ![]()
Изоклины горизонтальных касательных:
y=0 (ось абсцисс) и n0 – (1+x)(x+y) = 0 или: ![]()
Переменные x и y симметричны, поэтому изоклина вертикальных касательных
симметрична изоклине горизонтальных касательных.
Система имеет четыре особые точки:
1) x=0,
y=0 - неустойчивый
узел;
2) x=0, y=n0
- устойчивый узел;
3) x=n0 , y=0 - устойчивый узел;
4) и, наконец, симметричную точку - седло
![]()
В такой системе выживет один из видов: x или y. Его стационарная
концентрация определяется скоростью притока субстрата и экономическим
коэффициентом a. Как и в предыдущей системе (7.4) здесь причина отбора – неустойчивость симметричного состояния.
Генетический триггер Жакоба и Моно
Рассмотрим модель биохимической регуляции белкового синтеза, предложенную Жакобом и Моно в 1964 г. и математически разработанную Д.С. Чернавским в 1967 г. Эта модель показывает принципиальные возможности триггерных систем. Она легла в основу целой серии более подробных и конкретных моделей. Подробный вывод модели описан в книге Романовский Ю.М., Степанова Н.В., Чернавский Д.С. «Математическое моделирование в биофизике. М., 1975.
Схема взаимной регуляции двух систем синтеза ферментов изображена на рис. 7.9. Ген-регулятор каждой системы синтезирует неактивный репрессор. Этот репрессор, соединяясь с продуктом противоположной системы, образует активный комплекс, который обратимо реагируя с участком структурного гена, называемым опероном, блокирует синтез mRNK. Таким образом, продукт первой системы P1 является корепрессором второй системы, а продукт второй системы P2 - корепрессором первой. При этом в процессе корепрессии могут принимать участие одна, две и более молекул продукта.
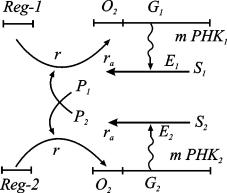
Рис. 7.9. Схема синтеза двух ферментов по Жакобу и Моно
Очевидно, что при таком характере взаимодействий при интенсивной работе первой системы вторая заблокирована, и наоборот. Простейшая система уравнений, описывающая такой тип взаимодействий, имеет вид
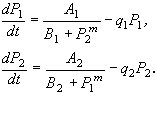
Здесь P1, P2 - концентрации продуктов, величины A1, A2, B1, B2, выражаются через параметры своих систем. Показатель
степени m показывает,
сколько молекул активного репрессора (соединений молекул продукта с молекулами
неактивного репрессора, который предполагается в избытке) соединяются с опероном
для блокировки синтеза mRNK.
Введем безразмерные переменные:

Опустив штрих у времени, перепишем систему в безразмерном виде:
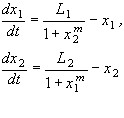
Исследование системы (7.19) показало, что при m = 0 фазовый портрет имеет одну устойчивую особую точку в
первом квадранте фазовой плоскости (рис. 7.10а) и не может описывать процессов переключения в системе.

Рис. 7.10. Главные изоклины на фазовой
плоскости системы 7.19. При m = 1. система имеет единственное
устойчивое стационарное состояние (а).
При m = 2 в системе три стационарных
состояния, два из которых (а и с) – устойчивые узлы, а третье (b)
– седло.
При m ³ 2 и определенных значениях отношения L1 / L2 > g система приобретает триггерные свойства. На фазовой
плоскости такая система имеет две устойчивые особые точки, между которыми
расположено седло (рис. 7.10 б).
Значение параметра g является бифуркационным, причем бифуркация имеет
триггерный характер (образуется седло). Отношение L1/L2 служит управляющим параметром, изменение значения L1/L2>g которого может
привести к смене стационарного режима в системе, как это было описано выше при
рассмотрении параметрического способа переключения системы. Величина параметров
L1, L2 зависит от многих биохимических характеристик: скорости
снабжения субстратами, активности ферментов, времени жизни ферментов, mRNK и продуктов и проч.
Фазовый портрет триггерной системы 7.19 приведен на рис. 7.11
Рис. 7.11. Фазовый портрет триггерной системы 7.19. Значения
параметров:

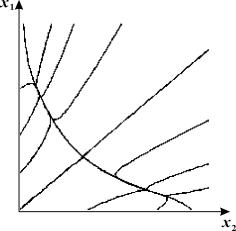
Таким образом, триггерные модели могут описывать процессы
отбора и дифференцировки. Подобные механизмы взаимодействия в распределенной системе
(при учете пространственной неоднородности и процессов переноса) могут
описывать процессы морфогенеза (формообразования). Эту модель – «распределенный генетический триггер» ‑ мы рассмотрим во второй
части лекций.
Литература
Романовский
Ю.М., Степанова Н.В., Чернавский Д.С. «Математическое моделирование в
биофизике. М., 1975.
Шноль С.Э. Физико-химические факторы биологической
эволюции. М., 1979
Воронцов
Н.Н. Развитие эволюционных идей в биологии. М.,1999
