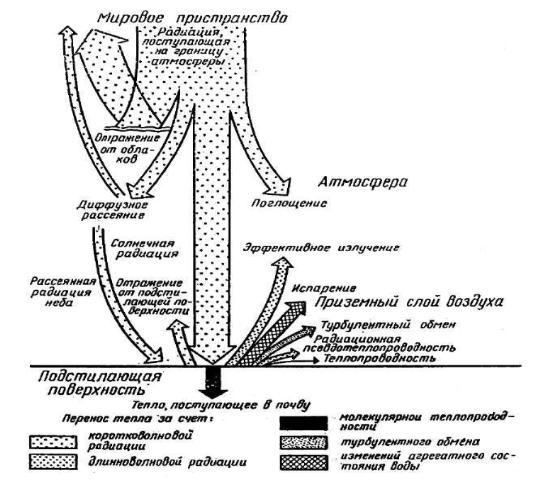
Рисунок 1 – Солнечная радиация и теплообмен у поверхности земли в летний полдень
FORESTRY - ЛЕСОВЕДЕНИЕМаркова Ирина Сергеевна |
Лесоведение является естественно – исторической основой практического лесоводства, его составной частью. Значение лесоведения в настоящее время расширяется в связи с государственным и общественным признанием многосторонней роли леса – объекта этой науки.
Важнейшие проблемы лесного хозяйства могут быть успешно решены только на основе знания природы леса. Поэтому лесоведение – наука о природе леса имеет большое значение для научно-технического прогресса лесного хозяйства.
Лесоведение как наука создана Г.Ф. Морозовым (1867-1920) в начале 20 века. Его предметом являются леса как природное единство всех составляющих его организмов и условий их обитания. Лесоведение дает классификацию леса по лесорастительным подзонам и типам леса, изучает закономерности возобновления древесных пород, их взаимоотношения, рост
и развитие леса, и смену состава древостоев с учетом хозяйственной деятельности человека.
Лесоведение как естественноисторическая часть лесоводства, раскрывая природу леса, рассматривает вопросы биологии и экологии леса в целом и составляющих его компонентов, географические аспекты леса, изучает закономерности в процессах возобновления и формирования леса, изменения характера леса в пространстве и во времени.
Основная задача лесоведения – изучать внутренние и внешние связи лесных биогеоценозов, их состав, продуктивность, динамику, потоки веществ
и энергии.
Лес – самая активная часть биосферы, лесной полог – преобразователь солнечной энергии, хранитель множества живых существ, и в этом состоит связь лесоведения с наукой о биосфере. Лесоведение широко использует все достижения лесной биогеоценологии и в целом науки о биосфере для совершенствования лесовыращивания так как ответственность лесоводов за состояние биосферы огромна.
Лесоведение или лесную биогеоценологию рассматривают как одну из естественных наук.
Лесоведение является сводом знаний, нескольких лесохозяйственных дисциплин и ряда общих наук, на которые опирается лесоводство. В этом смысле лесоведение включает дендрологию и физиологию древесных пород, лесную метеорологию и почвоведение, фитопатологию и энтомологию, отдел таксации леса, касающийся закономерностей в строении древостоев, биохимию лесных растений, лесную генетику, теоретическую часть лесной пирологии, учение о древесине и других продуктах леса (лесное ресурсоведение) и др.
Лесоведение имеет большое значение для практического лесоводства. Лесоводство занимается выращиванием леса для получения древесины и других продуктов леса и использования его с защитными, водорегулирующими, целебно-оздоровительными, эстетическими целями. Лесоводство разрабатывает методы повышения продуктивности леса и пути его улучшения, теорию и практику рубок леса в целях использования, возобновления и улучшения леса.
Для решения этих задач необходимо значение науки о природе леса, то есть лесоведения. Все вопросы современного лесоведения можно объединить в следующие основные разделы:
Значение леса в жизни человека продолжает увеличиваться. Постоянно расширяются потребности в древесине.
России принадлежит около 1/5 площади (1182 млн. га) всех лесов мира и 1/4 часть их запасов (82 млрд. кубометров). Преобладающими являются хвойные породы, запасы которых составляют 46 % от их мировых ресурсов.
По данным государственных учетов лесного фонда за 1966-1998 гг., лесистость страны увеличилась с 41 до 45%., а по обеспеченности стволовой древесиной на одного жителя она занимает место во втором десятке государств, уступая Канаде и Финляндии. Однако при таком богатстве народное хозяйство испытывает недостаток в древесине, так как наиболее распространены древостои лиственницы (43%), древесина которой до сих пор не находит широкого применения. Площадь под ценными породами составляет: дуб – 1 % площади; кедр – 6 %; ель – 12 %; сосна – 18 %.
В настоящее время 1/3 часть лесов недоступна для эксплуатации. 2/3 потребности в древесине приходится на европейскую часть России, где живет около 75% населения, а спелых лесов мало – 18 % от всех древесных запасов. Доставка древесины из северных районов очень дорога.
Все леса в зависимости от их хозяйственного значения, месторасположения и выполняемых функций разделены на 3 группы.
К первой группе из них отнесены леса, выполняющие средообразующие, защитные, санитарно-гигиенические и оздоровительные функции. Они выделены по всей территории страны и составляют 21% от площади всех лесов, а в европейской части 1/4 ее часть. Наибольшую долю в лесах I группы составляют запретные (водоохранные) полосы по берегам рек, озер и других водоемов. Они выполняют берегозащитные, водорегулирующие и водоочищающие функции.
Значительную территорию занимают климаторегулирующие и ландшафтно-защитные притундровые леса. Большую площадь имеют зеленые зоны вокруг городов, промышленных предприятий. В категорию защитных лесов входят полосы вдоль железных и крупных шоссейных дорог, противоэрозионные полезащитные леса, в том числе лесополосы, ленточные боры и степные колки.
Ко второй группе (7%) относятся леса, расположенные в густонаселенных районах с развитой транспортной сетью, а также леса в регионах с недостаточными лесными ресурсами. В европейской части их площадь близка к площади лесов первой группы, а в Сибири на Дальнем Востоке - мало. Леса этой группы также выполняют защитные функции, особенно водоохранные.
К третьей группе (72%) относятся леса многолесных районов, предназначенных для непрерывного удовлетворения потребностей народного хозяйства в древесине, но без ущерба для экологических функций этих лесов
Греческий философ Аристотель (384-322 г. до н. э.) предлагал изучать любое явление, начиная с истории его познания. Его интересовали проблемы леса, поскольку стихийная деятельность человека приводила к разрушению растительного покрова Греции. Леса, занимавшие более половины площади этой страны, вырубались, пастьба скота усиливала разрушение почвы водной эрозией, лес не возобновлялся. Этот процесс могли приостановить лесоводы, но в те века лесоводственные знания отсутствовали. В настоящее время лесистость Греции составляет 15-20 %, а на ранее обрабатываемых землях плодородие сохранилось только на 2 % площади.
Зачатки лесоводства возникли в лесах Древнего Рима. В переведенном на русский язык «Земледелие» Марка Порция Катона (234-149 гг. до н. э.) сообщается, что в виноградниках, на сельскохозяйственных полях или в специально отводимых местах высаживали иву, тополь, вяз, кипарис, сосну и другие древесные породы, руководствуясь их требовательностью к почве.
Луций Юний Колумелла в трактате о сельском хозяйстве в 55 г. н. э. излагает способы посадки леса, расширяет познания о лесоводственных свойствах древесных пород. Труды этого и других авторов обобщил Плиний Старший (23-79 гг н. э.). Он отметил, что леса, кроме того, что дают человеку древесину, выполняют почвозащитные и водоохранные функции.
Римские правила лесоводства действовали и в период Венецианской республики. Русский историк лесоводства Ф.К. Арнольд в 19 века сообщил, что, судя по дошедшим описаниям, в Северной Италии сохранялись весьма долго выборочные рубки почти в том самом виде, как было в Древнем Риме. Венеция в своих лесах в 15-16 столетиях вела прекрасное по тому времени лесное хозяйство. Леса были устроены, функционировало лесное управление, имелось учебное заведение, которое было подчинено Академии земледелия.
Россия в те времена была еще богата лесами. На карте 16 в. «Большой чертеж» леса простирались по берегам всех рек европейской части страны от истока до устья.
Период Российского средневековья (9-18 века) характеризуется все возрастающей эксплуатацией леса. Древесина широко использовалась на строительство судов, зданий, поэтому вдоль рек и вблизи населенных мест леса истощались.
Царь Алесей Михайлович (1629-1676 гг.) издал указы о строгом сбережении засечных лесов.
Петр 1 создал в 1719-1723 гг. лесное управление в виде коллегии адмиралтейства. По его приказу были описаны все леса вдоль больших и малых рек, там запрещалось вырубка ценных пород (дуб, ильмовые, клен и др.).
Для выращивания прочной корабельной древесины в те времена проводили обрезку сучьев. Передав металлургическим заводам Демидова на Урале большие площади лесов, Петр 1 составил правила ведения хозяйства в них, ввел систему лесосек и требовал заботиться о формировании на вырубках молодняков и об охране их от пожаров.
Первым отечественным ученым-лесоводом называют президента Академии наук А.А. Нартова (1737-1813), который описал лесокультурные приемы, выбор пород в зависимости от плодородия почвы, исследовал воздействие деревьев друг на друга и на живой напочвенный покров.
Большой вклад внес в развитие лесоводства известный ученый и литератор А.Т. Болотов (1738-1833). Рубки ухода он рассматривал как систему, предлагая пятикратный уход за хвойными породами.
Научными исследованиями лесов занимался М.В. Ломоносов (17111765), обративший внимание на их почвоулучшающую роль, воздушное питание растений и т.п.
В 18 веке лесами занимались известные географы В.Н. Татищев, И.Г. Гмелин, С.П. Крашенинников, Э. Лаксман, П.С. Паллас и др.
Лесоведение и лесоводство тесно связано. Лесоведение как учение о природе леса оформилось в начале 20 в. в недрах лесоводства благодаря Морозову и получило дальнейшее развитие во 2-й половине столетия. В развитии лесоведения большую роль сыграли (в середине 20 в.) идеи основоположника биогеоценологии советского ученого В.Н. Сукачева.
Значительный вклад в развитие лесоведения внесли также ученые Г.Н. Высоцкий, Л.А. Иванов, М.Е. Ткаченко, Н.В. Третьяков, А.А. Тюрин, П.С. Погребняк и др. Их ученики, а также последующие поколения исследователей.
Мелехов И.С. Лесоведение. М.: МГУЛ.- 2004, 398 с.
Дополнительная литература:
Погребняк П.С. Общее лесоводство. Колос. М.: 1968, 440 с.
В лесоведении термином «лес» обозначают элементарную единицу биосферы. Лес является и составной частью географического ландшафта – однородной в генетическом отношении территории, на которой наблюдается закономерное повторение участков, тождественных по геологическому строению, форме рельефа, гидрологии, микроклимата, почвам и фитоценозам.
Чтобы лучше понять, что такое «лес», необходимо сравнить деревья, растущие на свободе и в лесу, а также изучить взаимодействие тех и других со средой.
Дерево, выросшее в лесу, имеет более прямой ствол, большую высоту, но меньший диаметр по сравнению с деревьями, растущими на свободе.
Их толщина от комля к вершине уменьшается постепенно, и они более полнодревесны, чем деревья, выросшие на свободе.
Крона дерева, растущего в лесу, уже и сосредоточена в верхней части ствола. Крона дерева, растущего на свободе, располагается почти по всему стволу.
Не всякий участок земли с растущими на нем деревьями можно назвать лесом. Это может быть парк или вырубка с сохранившимся подростом.
Критерием, по достижении которого при смыкании крон в молодом возрасте начинается влияние деревьев друг на друга, является величина сомкнутости древесного полога 0,4. При сомкнутости древесного полога менее 0,4, территория относится к непокрытой лесом площади, более 0,4 относится к покрытой лесом, то есть рассматривается как лес, с присущим ему влиянием деревьев друг на друга и на другие компоненты биогеоценоза (микроклимат, почву и др.).
Лес является частью географического ландшафта. Это один из основных типов растительности Земли, сложное сочетание множества разнообразных взаимосвязанных организмов.
Принято считать, что лесная среда образуется под сомкнутым древесным пологом и на расстоянии от стены леса, равном 1-2 высотам деревьев.
К факторам лесообразования относятся:
Под внутренними, экологическими свойствами древесных пород понимается отношение видов к климату, почве и биологические свойства, долговечность, особенности роста, размножение и т.д.
Лесообразующее значение географической среды часто связано с экологическими свойствами древесных пород.
Биосоциальные отношения между растениями известны давно и закрепились в термине «породы-спутники» и выражении «осина-нянька ели». Но известны и примеры неблагоприятного воздействия.
Биосоциальные отношения между растениями и фауной наглядно выявляются при воздействии лося на сосну. Увеличение поголовья лосиного стада приводит к уничтожению сосновых молодняков.
Вмешательство человека повлияло на распространение березы и осины после сплошных рубок на северо-западе России, а в южной части способствовало смене семенных дубрав на порослевые. Отдыхая в лесу, выпасая скот, проводя рубки ухода, человек нарушает «подвижное равновесие», чем активизирует размножение вредителей и болезней в лесу.
Большую роль играют лесные пожары. Причины их разнообразны. От молнии ежегодно в мире возникает около 50000 лесных пожаров. В таких случаях огонь является самостоятельным фактором лесообразования. Лесные пожары способствуют смене темнохвойных лесов на светлохвойные. В районе воздействия огня в почву поступает большое количество зольных элементов, изменяется ее влажность, тепловой режим верхних слоев, увеличивается активность нитрофицирующих бактерий. Благодаря этому даже на песчаных свежих почвах тайги могут расти сосняки 1 класса бонитета.
Все эти факторы лесообразования создали в лесу различные насаждения, отличающиеся по тем или иным признакам. Элементарной естественноисторической единицей такого разнообразия считается лесной биогеоценоз.
Лесной биогеоценоз состоит из биоценоза, включающего все виды растений (фитоценоз), животный мир, населяющий лес: млекопитающие, птицы и другие позвоночные, а также насекомые, черви, моллюски и другие беспозвоночные (зооценоз) и микроскопические организмы в почве, в органах растений и животных (микробоценоз). Так как в процессе жизнедеятельности биоценоз изменяет почву, атмосферу в лесу, то в биоценоз включаются факторы мертвой среды, которые называются экотопом. Экотоп в этом случае состоит из атмосферы с ее лесными особенностями (климатоп) и почвы с горной породой и грунтовыми водами (эдафотоп). Все эти составные части влияют друг на друга и на жизнь лесного биогеоценоза. Лес не может существовать, если исключить из него какую-либо часть.
Основными компонентами леса являются древостой, подрост, подгон, подлесок, живой напочвенный покров и внеярусная растительность.
Лесное насаждение, которое представляет собой лесной фитоценоз, трактуется по ОСТ 56-108-98 как «совокупность растений, состоящих из древостоя, а также часто подроста, подлеска и живого напочвенного покрова, объединенных однородными лесорастительными условиями участка леса и характеризующаяся определенной внутренней структурой». Сюда же следует включать и внеярусную растительность, к которой относятся лианы, а также эпифиты (мхи, лишайники) и грибы на стволах деревьев.
Живой напочвенный покров – совокупность мхов, лишайников, травянистых растений, кустарничков и полукустарников, произрастающих на лесопокрытых и лесонепокрытых землях.
Подлесок – кустарники, реже деревья, произрастающие под пологом леса и не способные образовать древостой или войти в состав древостоя в конкретных лесорастительных условиях.
Подрост – молодое поколение древесных растений под пологом древостоя или на лесонепокрытых землях, способное образовать новый древостой.
Древостой – совокупность деревьев, иногда кустарников, являющаяся основным компонентом насаждения.
Характерными признаками древостоя являются: происхождение, форма, состав, возраст, полнота, бонитет, средний диаметр ствола, высота, густота, запас, и др.
По происхождению различают древостои семенные, которые возникли из семян, и вегетативные, включающие порослевое, корнеотпрысковое и отводковое возобновление леса.
Форма – совокупность ярусов насаждения. По форме насаждения могут быть простые – одноярусные и сложные – многоярусные.
По составу древостои подразделяются на чистые, состоящие из одной породы или с единичной примесью деревьев, и смешанные, состоящие из двух и более пород. Состав пород обычно обозначают по 10-бальной системе. Так 10С – обозначает чистый сосновый (10/10 сосны) древостой; 6С4Е+Ос указывает на смешанный древостой, так как помимо сосны, которой 6/10, здесь имеется еще 4/10 ели и единично (+) осины. В практике
количественно выражают коэффициентом, пропорциональным объему стволовой древесины, а в древостоях до 10 лет – по густоте или по соотношению сомкнутости крон древесных пород. Сумма коэффициентов всех пород древостоя должна составлять 10.
Порода, имеющая больший коэффициент, называется преобладающей. По хозяйственному значению древесные породы делят на главные, второстепенные и нежелательные. Главная древесная порода – древесная порода, которая в определенных лесорастительных и экономических условиях наилучшим образом отвечает хозяйственным и экономическим целям. Древесные породы меньшей хозяйственной ценности, чем главная, относятся к второстепенным, а породы, которые не нужны в определенных экономических условиях, - к нежелательным.
По возрастному строению различают одновозрастные и разновозрастные древостои. В одновозрастных древостоях почти все деревья находятся в пределах одного класса возраста (5-10-20 или 40 лет). Если в разновозрастных древостоях есть более 15% деревьев, выходящих за эти пределы, их относят к другому классу возраста и выделяют как самостоятельные поколения.
Полнота древостоев характеризует степень заполнения древостоем пространства в горизонтальной плоскости определенной территории, выражаемая суммой поперечных сечений стволов составляющих древостой деревьев (ОСТ 56-108-98).
Выделяется полнота «абсолютная», выражаемая в абсолютных единицах на единицу площади – квадратных метрах на гектар и «относительная» - в десятых долях от единицы, соответствующей эталонной полноте древостоя в данных условиях.
Сомкнутость древостоя – степень заполнения пространства участка леса кронами деревьев, определяемая по их общей проекции на горизонтальную плоскость.
Сомкнутость полога древостоя выражается в десятых долях от единицы, соответствующей полной сомкнутости.
Древостой с сомкнутостью полога, обеспечивающей формирование и сохранение лесной среды, является сомкнутым.
К сомкнутым относятся обычно древостои с сомкнутостью или полнотой 0,3 и выше, молодняки – 0,4 и выше.
По сложению различают горизонтальную, вертикальную и ступенчатую сомкнутость полога.
Густота – число стволов на 1 га.
Запас древостоя – общее количество древесины стволов растущих деревьев древостоя.
Бонитет – отражает чаще всего потенциальную производительность древостоя и обусловлен лесорастительными условиями и требовательностью древесной породы к почве.
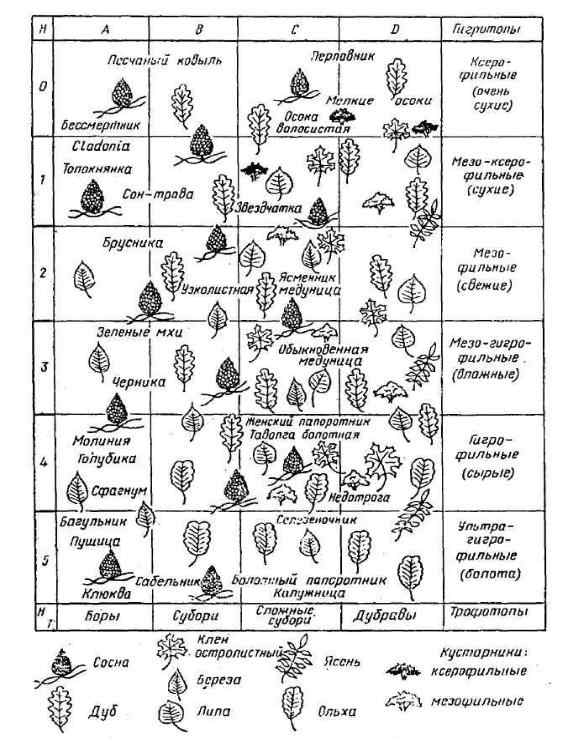
Лесорастительные условия – это комплекс климатических, гидрологических и почвенных факторов.
Тип леса – лесоводственная классификационная единица, объединяющая леса с однородными лесорастительными условиями определенного типа, соответствующим им породным составом древостоя, другой растительностью и фауной.
Лесной биогеоценоз внутри себя неоднороден. Так как это динамическая система, то в ней имеются остатки прошлого типа биогеоценоза и зачатки новых. Под кронами деревьев различных пород видовой состав напочвенного покрова отличается. Разница существует и в составе почв, который зависит от полноты древостоя.
В «окнах» древесного полога другой микроклимат, а групповой подрост отражает многообразие всех компонентов леса на этом месте. Такие более мелкие структурные образования биогеоценоза, отличающиеся по составу пород, полноте, почве, особенностями микрофлоры Н.В. Дылис назвал биогеоценотическими парцеллами.
Частью парцеллы является синузия. Н.В. Сукачев понимал синузии как структурные части фитоценоза, характеризующиеся определенной экологией составляющих их видов и пространственной обособленностью, а следовательно, и особой фитоценотической средой, создаваемой растениями данной синузии.
Каждая синузия занимает определенную экологическую нишу, ограничена в пространстве и во времени. В лесных фитоценозах, образованных несколькими ярусами и многими видами растений можно выделить десятки синузий. Наибольшее значение имеют синузии древесного яруса, подлеска.
Лесной биогеоценоз можно считать открытой кибернетической системой, так как он обладает свойствами саморегуляции. Регуляция находит законченное выражение в том, что в сходных условиях среды могут формироваться однотипные биогеоценозы. Примером саморегуляции экосистемы может быть разрастание подлеска при снижении полноты древостоя.
Экосистема включает в себя подсистемы на уровне компонентов леса или их частей, которым также свойственна саморегуляция. Влияние саморегуляции можно проследить на примере густоты девственного разновозрастного древостоя. После гибели крупного дерева на его месте появляется много молодых особей. Но густота древостоя на определенной площади (0,5-2,0 га) повышается ненадолго, так как кроме старого дерева в порядке естественного изреживания погибает много нестарых, что свидетельствует о гомеостазе системы (Гомеостаз – способность системы организмов поддерживать динамическое равновесие в изменяющихся условиях среды).
Различают резистентную и упругую устойчивость экосистемы. Резистентная – это способность экосистемы сопротивляться пертурбациям (нарушениям), поддерживая неизменной свою структуру и функцию. Упругая устойчивость – это способность системы восстанавливаться после того, как ее структура и функции были нарушены. Саморегуляция лесного биогеоценоза обеспечивается и тем и другим видом устойчивости.
Резистентная устойчивость позволяет, например, почве сопротивляться интоксикации путем:
И только при неограниченном поступлении загрязняющих веществ сильная интоксикация вызывает необратимое разрушение лесного биогеоценоза.
Упругая устойчивость экосистемы позволяет осуществлять рубки ухода, внедряться в лес с техникой. Через несколько лет древостой снова смыкается.
Таким образом, благодаря саморегуляции экосистемы леса прирост массы отдельных компонентов подчинен функциональному целому, и популяции не растут безгранично. Однако и саморегулирующиеся биогеоценозы подвергаются стрессам от засух, загрязнения воздуха и т. д.
Для повышения устойчивости в экосистему надо вводить различные виды лесной древесной и кустарниковой растительности, обладающей высокой толерантностью (то есть способностью выживать при большом разбросе параметров факторов среды) и создающие синузии; при рубках ухода оставлять стволы различной величины, не нарушать биогруппы, регулировать плотность животных, восстанавливать плодородие почв и т. п.
Лесные биогеоценозы, имея черты гомеостатической системы, одновременно являются и адаптивной системой, т. е. изменяются в направлении повышения устойчивости к изменившимся условиям среды.
На лесной биогеоценоз из абиотических факторов действуют свет, тепло (холод), осадки (жидкие, твердые), ветер, составные части воздуха (СО2, промышленные газы, пыль, радионуклиды), атмосферное электричество и т. д.
Многие абиотические факторы резко влияют на биологическую производительность лесного биогеоценоза, а при сильном воздействии вызывают гибель насаждения.
1. Дайте определение леса как природного явления.
2. Перечислите основные компоненты и признаки леса.
3.Охарактеризуйте факторы лесообразования и их значение для современного лесоводства.
4. Расскажите о структуре лесного биогеоценоза.
5.Приведите доказательства того, что лес биологическая саморегулирующаяся система.
Основная литература:
Мелехов И.С. Лесоведение. М.: МГУЛ.- 2004, 398 с.
Дополнительная литература:
Погребняк П.С. Общее лесоводство. Колос. М.: 1968, 440 с.
1 Общие сведения об экологии леса
2 Лес и климат. Климат и распространение лесов
3 Солнечная радиация и лес
4 Разнообразие лесов на земном шаре
5Лесорастительные подзоны России
6 Вертикальная зональность
7 Лесорастительное и лесохозяйственное районирование
Лесная экология изучает популяции и территориальные совокупности видов – лесной биогеоценоз.
Лесной биогеоценоз имеет не только внутренние связи между компонентами, но и внешние, так как происходит обмен веществом и энергией с другими биогеоценозами, с атмосферой, литосферой, космосом.
Экологические факторы подразделяются на:
1)климатические – или атмосферные факторы – свет, тепло, осадки, состав, влажность, движение воздуха (ветер), испарение и др.;
2)эдафо-орографические – почвообразующие породы, эрозионные и наносные процессы, физические свойства почвы, ее химический состав, концентрация почвенного раствора, состав почвенного воздуха, мощь корнедоступной толщи, грунтовые воды, рельеф;
3)биотические – птицы, насекомые, другие животные, бактерии, грибы и высшие растения;
4)антропогенные – действие человека, выражающееся в рубках, лесоосушении, пастьбе скота, загрязнении
атмосферы и т. д.
Экологические факторы действуют на все организмы, в том числе на деревья комплексно. Роль каждого фактора повышается, если потребность в нем удовлетворяется не полностью.
Жизнь древостоя необходимо рассматривать с экологической точки зрения.
Климат оказывает влияние на лес. В различных климатических регионах различен состав насаждений, их продуктивность. Не только климат влияет на лес, но и лес влияет на климат.
Г.Н. Высоцкий придавал большое значение отношению величины осадков к величине испарения, или коэффициенту увлажнения: при коэффициенте больше 1 – климат влажный, свойственный лесной зоне; при 1
– соответствует лесостепи; коэффициент 0,5 – характерен для полупустыни. Г.Г. Селянинов (1933) ввел показатели теплообеспеченности и
влагообеспеченности. Первый характеризуется суммами активных температур (выше 100С). Показатель влагообеспеченности дан в виде гидротермического коэффициента, представляющего отношение осадков за вегетационный период к сумме температур за этот же период, уменьшенное в 10 раз.
Шведский биоклиматолог С. Патерсон (1956) сделал попытку установления связи потенциальной продуктивности леса (прироста древесины) с основными климатическими факторами в виде индекса CVP ( С
– климат, V – растительности, Р – продуктивность).
CVP = Т/Та · Р · G/12 · Е/200;
Где Т – среднемесячная температура наиболее жаркого месяца, 0С;
Та – разница между среднемесячными температурами наиболее жаркого и наиболее холодного месяца;
Р – среднегодовое количество осадков, мм;
G – продолжительность вегетационного периода, мес.;
Е – местная солнечная радиация (обусловливающая транспирацию). Выявив связь CVP – индексов и прироста древесины для 1426 точек, С.
Патерсон нанес на карту мира кривые с одинаковыми показателями и выделил 6 зон потенциальной продуктивности.
Основные климатические факторы (недостаток тепла и влаги), ограничивающие распространение леса, влияют на производительность древостоя. К.Б. Лосницкий рассчитал потенциальную продуктивность древесных пород европейской части в зависимости от суммы активных температур свыше + 100С и наибольший потенциал установил для ели. Выявлены связи классов бонитета древостоев различных пород с суммой активных температур 350-12500 в горах Южной Сибири. Количество осадков (600-1000 мм) в значительной степени сказалось на производительности кедрачей.
Биологическая продуктивность (рост деревьев, формирование годичных колец) тесно связанна с продолжительностью вегетационного периода, соотношением тепла и влаги.
Согласно климатической классификации на земном шаре выделяют следующие типы климата:
- климат тундры со средней суточной температурой самого теплого месяца от 0 до 10-120С и типичной низкорослой растительностью и осадками менее 300 в год;
- климат тайги – с температурой самого теплого месяца от 10 до 200С и осадками 300-600 мм в год с преобладанием их в летнее время, здесь произрастаю хвойные и лиственные леса;
-климат смешанных хвойных и лиственных лесов умеренной зоны со средней температурой 4-х теплых месяцев от 10 до 22 0С и осадками 400-700 мм в год;
-муссонный климат умеренных широт с малоснежной зимой и выпадением осадков в теплое время года (85-90% годовой суммы осадков);
-климат степей с температурой летних месяцев 20-330С и количеством осадков 200-400 мм, преобладает степная растительность;
- климат земноморский с жарким и сухим летом, теплой и влажной зимой;
-климат зоны субтропических лесов с температурой самого холодного месяца выше 20С, с большим количеством осадков.
Кроме этих типов климата, выделяют климат внутриматериковых пустынь умеренного пояса, субтропических пустынь, саванн, или тропической лесостепи и влажных тропических лесов.
Различным типам климата соответствуют особые леса.
С особенностями климата связаны сроки посева и посадки леса, способы обработки почвы. Климат, метеорологические условия определяют степень опасности возникновения и распространения лесных пожаров. Поэтому в практике охраны лесов от пожаров широко используют данные метеослужбы. С климатом связаны сезонные особенности рубок леса, организация лесозаготовок. Климат обуславливает сроки атмосферной сушки лесоматериалов и многие другие стороны лесного производства.
Климат понятие интегральное и его влияние на лес сказывается комплексно, в разных взаимосвязанных составляющих его элементов. Свет, тепло, осадки, влажность, состав и движение воздуха – все эти взаимосвязанные элементы климата оказывают большое влияние на лес.
В начале 20 века немецкий естествоиспытатель А. Гумбольд внес значительный вклад в выявление закономерностей распространения растительности в связи с климатом. Он ввел понятие изотермы – линии, соединяющей точки одинаковых температур на географической карте. Если
проследить за северной границей лесов, то она совпадает с июльской изотермой + 100С. Значит, основным ограничивающим фактором распространения лесов на север является недостаток тепла, особенно в почве. Он ограничивает не только рост вегетативных органов, но и семеношение. В лесах на северной границе семена созревают редко, иногда раз в столетие.
ВАзиатской части России на распространение лесов кроме недостатка тепла влияют и зимне-весенние условия: сухость воздуха и ветры, вызывающие потерю влаги растениями. Эти факторы отражаются на составе лесов.
По мере потепления климата в первой половине столетия наблюдалось продвижение древесной растительности на север со скоростью 20-150 м в год. Это был короткий цикл, не изменивший многовекового похолодания. Как показал радиоуглеродный метод определения возраста ископаемой древесины 518 ± 150 лет назад, на безлесном сейчас полуострове Таймыр произрастали леса. Это был период потепления климата 8-4 тыс. лет назад.
Еще теплее было в каменноугольный и третичный периоды, когда в эоценовую эпоху (35-55 млн. лет назад) современная Центральная Европа находилась примерно на 5-100 севернее экватора, а на Шпицбергене росли мамонтовые деревья и болотные кипарисы. Затем перемещение полюсов вызвало сначала медленное, затем быстрое похолодание. Изучение окаменелостей древесины, пыльцы и спор в отложениях болот и озер показало, что во время оледенения леса отступали к побережьям Средиземного и Черного морей и восточнее – к южному берегу Каспия.
В межледниковые периоды продолжительностью 40000 до 150000 лет в Центральное Европе наблюдались характерные циклы смены растительности: тундровые, березовые, березо-сосновые, дубовые, дубовограбовые, грабово-буковые, еловые и пихтовые леса, сосново-еловые, сосново-березовые, тундра.
|
Рисунок 1 – Солнечная радиация и теплообмен у поверхности земли в летний полдень |
Южная граница распространения лесов также связана с климатом и совпадает с величиной отношения количества испарившейся влаги с водной поверхности к количеству осадков.
Роль относительного увлажнения установлена и для нижней границы лесов в горах Южной Сибири.
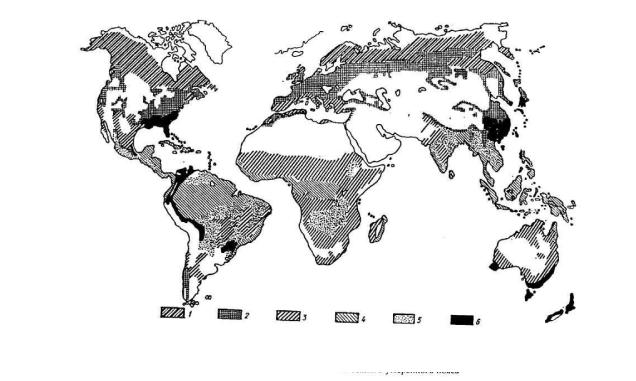
Исходным климатообразующим началом, обуславливающим характер климата и его элементов, в первую очередь света и тепла, является энергия Солнца, солнечная радиация.
На разных географических широтах, в разное время года и суток интенсивность солнечной радиации неодинакова вследствие различных углов падения солнечных лучей. Не вся радиация достигает поверхности Земли. Большое влияние на нее оказывает атмосфера (рисунок 1).
Солнечная радиация оказывает большое повседневное влияние на жизненные процессы леса, обуславливает фотосинтез, транспирацию, тепло
и влагообмен и т. д. Еще есть одна особенность проявления солнечной энергии – это многолетние циклические изменения уровня солнечной активности, повторяющиеся через несколько лет.
Влияние солнечной активности может осуществляться и через воздействие геомагнитных полей Земли, ионизацию атмосферы и т. п. Вызванные подобными воздействиями изменения леса проявляются поразному в разных регионах, на разных почвах, способствуя повышению прироста в одних случаях, усыханию деревьев в других.
Образующиеся вследствие повышенной солнечной активности длительные сухие периоды повышают опасность возникновения лесных пожаров.
Влияние солнечной радиации на лес велико и проявляется в разных формах. Благодаря этому влиянию лес может быть индикатором изменения солнечной активности и климата нашей планеты. В свою очередь лес сам оказывает влияние на поступающую радиацию. Лес – один из наиболее сильно и многосторонне воздействующих на поступающую солнечную радиацию видов подстилающей поверхности на Земле. Он отличается малой отражательной способностью и высоким радиационным балансом.
Лучистая энергия Солнца, попадая на поверхность полога леса, частично отражается, частично поглощается деревьями и др. растениями, часть ее через просветы в пологе проникает вглубь лесного сообщества и доходит до почвы. При этом происходят количественные и качественные изменения этой энергии. Радиация под пологом леса ослабляется и меняет свой спектральный состав.
Радиационный баланс леса выше радиационного баланса других видов поверхности земли. В целом уравнение радиационного баланса выглядит следующим образом:
В = S + q+ Ea – (α + E3), где
В – радиационный баланс леса;
S – прямая радиация Солнца; q – рассеянная радиация;
Еа – тепловое противоизлучение атмосферы; α – отражение солнечного излучения от подстилающей поверхности;
Е3 – собственное тепловое излучение земной поверхности.
Полог аккумулирует летом 20-45 % радиации или на 15-20% больше, чем открытое пространство. Эта способность радиации в лесу за вегетационный период распределяется по формуле теплового баланса:
В = Ф + ИФ + Т + Р + А Где: Ф – расход тепла на фотосинтез;
ИФ – расход на физическое испарение осадков, задержанных кронами, травостоем и воды с поверхности почвы (10-30%);
Т- расход на транспирацию (40-60%); Р – расход тепла на турбулентный воздухообмен между лесом и
открытым пространством или проходящими воздушными массами (10-50%);
А – аккумуляция тепла в почве и наземной биомассе и в среднем за год равна 0 (меньше 5%).
Основной расход солнечного тепла у деревьев приходится на транспирацию, так как листовой индекс у них выше, чем на лугу, вырубке или болоте.
Покрытая лесом площадь планеты составляет около 3,8 млд. га, или 29,7 % земной суши. Примерно половина (53%) лесов произрастает в умеренной зоне, остальные в тропической; сомкнутые древостои составляют соответственно 80 и 69%. Схематическая карта лесов мира приведена на рисунке 2.
|
Рисунок 2 – Схематическая карта лесов мира: 1 – хвойные леса холодной зоны; 2 – смешанные леса умеренного пояса; |
В северном полушарии граничат с тундрой северные леса холодной (бореальной) зоны, которые в России называются термином тайга. Хвойные породы вечнозеленым покровом аккумулируют больше солнечной энергии и имеют более длительный период фотосинтеза (с первой оттепели и до глубокой осени).
Эта самая крупная растительная зона земного шара, не имеющая аналогов в южном полушарии. Состав пород: из хвойных – ель, сосна, пихта, лиственница; из мелколиственных – береза, тополь, ольха, ива.
Хвойные леса бореальной зоны имеют большое климаторегулирующее значение и являются источником деловой древесины: 70% древесины хвойных в мире заготавливается в лесах этой зоны. Южная граница зоны хвойных лесов совпадает в Америке и Европе с северной границей ареала клена, дуба, в Сибири примыкает к степям и полупустыням.
Южнее этой зоны простираются смешанные леса умеренной зоны, или летне-зеленые лиственные леса.
Они связаны с умеренно прохладным морским климатом. Их полоса сужается по направлению внутрь континента, а затем исчезает, уступая место прериям, степям и полупустыням.
Ближе к экватору в обоих полушариях вблизи морей и океанов формируется субтропический тип растительности с влажными лесами теплого климата. Они занимают промежуточное положение между летнезелеными лиственными и тропическими лесами. Типичными для этой зоны являются леса Колхиды в Грузии.
Естественные леса субтропиков истощены, низкопродуктивны и заменяются плантациями быстрорастущих древесных пород.
В экваториальной зоне по берегам океанов произрастают дождевые леса с вечнозелеными широколиственными породами. В них число видов на 1 га может достигать 100, а эксплуатационное значение нередко имеют один или несколько стволов.
Дождевые леса сосредоточены в трех районах:
1 в северной части Южной Америки (бассейн реки Амазонки); 2 в западной Экваториальной Африке (бассейн р. Конго);
3 в Индо-Малакском регионе, включающем западное побережье Индии, большую часть полуострова Индокитай и северо-восточное побережье Австралии.
Вглубине континентов за пределами дождевых лесов в условиях экваториальной зоны при слабом воздействии морских ветров и количестве годовых осадков 100-2000 мм наблюдается сухой период продолжительностью месяц и более во время которого большинство деревьев сбрасывает листву. Такие насаждения называются листопадными лесами.
При меньшем выпадении осадков образуются сухие леса или саванны. Они занимают часть тропической Африки, тропической Австралии и распространены к северу и югу от бассейна Амазонки. Тропические леса являются достоянием развивающихся стран, которые 82 % заготовленной древесины расходуют на топливо.
Лесной фонд России составляет 1,2 млрд. га. Леса произрастают в различных климатических условиях. При среднем приросте стволовой древесины на 1 га покрытой лесом площади 1,32 м3 в одних регионах он составляет менее 1 м3, в других более 3м3, что свидетельствует о неоднородности лесов.
Суть растительного и животного мира с климатом была выявлена в 18 веке. Первый опыт зонирования лесов принадлежит Г.И. Танфильеву.
В 1960 г. институт леса АН СССР разбил лесорастительную географическую зону на ряд лесорастительных подзон: северную, среднюю и южную тайгу, подзону хвойно-широколиственных лесов (смешанных) и подзону широколиственных лесов.
В1980 г. данные геоботанической карты были обновлены и в настоящее время подзона смешанных лесов до Урала называется широколистенно-еловой, а в широколиственной подзоне выделена полоса широколиственных лесов с елью. В настоящее время отдельно рассматриваются горные леса Урала, равнинные леса Западной Сибири, горные леса Сибири, горные леса Дальнего Востока и другие леса регионов.
Северо-таежная европейская подзона вытянута полосой шириной от
250км на востоке до 600 км на западе. Северная граница проходит посреди
Кольского полуострова и далее совпадает с Северным полярным кругом. Южная граница совпадает с северной широтой 630. Подзона характеризуется
низкопродуктивными еловыми, сосновыми и березовыми лесами 4 и 5 классов бонитета с приростом менее 1 м3 на 1 га.
Подзона северной тайги занимает площадь в виде 400-450 км полосы, резко сужающейся в Карелии. Леса более сомкнуты и продуктивны,
древостои в основном 3 – 4 классов бонитета, имеющие средний прирост около 1,5 м3 на 1 га. Наряду с древесными породами, свойственным тайге, здесь появляется ольха черная и липа мелколистная. Главная ценность средней тайги – еловые леса, в которых заготавливается самая плотная древесина. Однако проведение сплошных концентрированных рубок приводит к смене ели березой и осиной.
В подзоне южной тайги, протянувшейся в виде узкой полосы шириной
200-300 км с южной границей вблизи городов Нарва, Псков, Новгород, Ярославль и далее по северной широте 570, преобладает ель. Но сохранились остатки широколиственных лесов из дуба черешчатого, клена остролистного,
ясеня обыкновенного, вязов гладкого и шершавого. Ельники I-III классов бонитета. Средний прирост стволовой древесины в лесах составляет 2,0 м3 на 1 га.
В этой подзоне развиты лесозаготовки, на больших площадях создаются культуры ели и сосны.
Для европейской подзоны хвойно-широколиственных лесов характерно одинаковое распространение как хвойных, так и лиственных пород: осины, березы, дуба, липы, клена, вяза, ясеня и на юго-западе граба.
Леса этой подзоны отличаются высокой производительностью: средний прирост 3,0 м3 на 1 га.
Лесорастительная подзона широколиственных лесов выражена в югозападной части лесной зоны. Здесь осадков выпадает 600 мм в год.
Характерным для широколиственных лесов является большое разнообразие древесных и кустарниковых пород.
Леса этой подзоны выполняют разнообразные функции: почвозащитные, рекреационные, водо-очищающие и др.
На востоке тайги продолжается полосой подзона горных лесов Урала, характеризующаяся разнообразием лесорастительных условий из-за большой протяженности в меридиональном направлении, вертикальной зональности и различного количества осадков на западном и восточном склонах хребта. Общим для этих лесов являются их высокое почвозащитное и водорегулирующее значение. Здесь преобладают хвойные виды: ель сибирская, пихта сибирская, лиственница сибирская, сосна обыкновенная и сибирская, береза и осина.
За последние 40 лет лесистость и средний прирост снизились с 40% и 2,0 м3/га и 1,4 м3/га.
Подзона равнинных лесов Западной Сибири представлена сибирскими видами ели, пихты, лиственницы и сосны (кедр). На песках и болотах распространена сосна обыкновенная, а на границе со степью преобладает береза, изредка липа, вяз. Это край болот. Лесистость составляет 33 %, а из-за заболоченности почв средний прирост древесины находится на уровне 1,6 м3 на 1 га.
За Енисеем простираются горные леса Сибири. Эта территория подразделяется на Среднюю Сибирь, Восточную Сибирь (собственно горная страна) и горы Южной Сибири в пределах которых выделены 2 лесорастительные подзоны: подзона лесов Восточной Сибири и подзона горных лесов Южной Сибири. В этом районе сосредоточена треть запасов спелых древостоев РФ. Однако основной породой является лиственница Гмелина, древесина которой применения не нашла.
Второе место по площади занимают сосняки (сосна обыкновенная). На третьем месте – кедровики из кедровой сибирской сосны.
Большая территория Восточной Сибири занята низкопродуктивными древостоями хвойных пород.
Основная проблема лесоводства в горных лесах Восточной Сибири сводится к пересмотру принципов ведения рубок и предупреждение и борьба с лесными пожарами.
Эти же проблемы типичны и для горных лесов Дальнего Востока, представленных в основном подзоной дальневосточной тайги. На территории Дальнего Востока наблюдается разнообразие климата от океанического до континентального, преобладает сезонная и многолетняя мерзлота.
На юге Дальнего Востока представлены другие лесорастительные подзоны – хвойно-широколиственные леса Амурско-Уссуррийской провинции и широколиственные леса бассейна Уссури. Из хвойных пород распространенные кедр корейский, лиственница амурская, пихта цельнолистная, ели аянская, сибирская и корейская, редко тис; из лиственных
– береза даурская, железная, желтая, каменная, дуб монгольский, липа амурская и манчжурская, бархат амурский, орех манчжурский, ильм японский и др.
Островные и пойменные леса встречаются в степной природной зоне. Искусственными являются леса Донлесхоза Ростовской области, Среднечелбасскаго в Краснодарском крае и др. Примечательны естественные ленточные боры в западной Сибири. В степи созданы полезащитные лесные полосы.
Кроме горизонтальной (широтной) зональности в распространении растительности в горах наблюдается вертикальная зональность, или высотные пояса растительности.
С подъемом относительно уровня моря понижаются атмосферное давление и температура воздуха. При адиабатическом процессе с маловлажным воздухом через каждые 100 м высоты температура понижается на 10. Если такой воздух, встретивший препятствие в виде возвышенности или горы, поднимается вверх, то температура его понижается и увеличивается относительная влажность. При влажности воздуха 100 % начинается конденсация паров с выделением тепла, и выпадают осадки. Адиабатический процесс переходит в политропический, то есть температура с подъемом на каждые 100 м падает не на 1 градус, а на 0,5-0,60. Следовательно, чем выше поднимаешься в гору, тем холоднее и влажнее становится климат.
Особенно резко возрастает количество осадков на склонах, обращенных к господствующим ветрам.
Перевалив через Уральский хребет, воздушные массы опускаются, происходит адиабатический процесс сжатия воздуха, он нагревается после каждых 100 м на 10 на западных склонах Урала распространены ельники и пихтачи, а на восточных – древостой их ксерофитной сосны обыкновенной.
Такое изменение климата в горах влияет на растительность, а вместе с нею и на почвообразование. Так, на северо-восточных макро склонах Карпат нижний пояс представлен дубравами. На высоте 900-1200 м над уровнем моря расположен «подпояс» биологически устойчивых смешанных насаждений с елью и пихтой в первом ярусе и с буком, явором и ильмом во втором на бурых мощных почвах. Ельники здесь имеют максимальную продуктивность. На высоте 1200-1700 м над уровнем моря простирается «подпояс» чистых ельников III-Y классов бонитета с маломощными бурыми горно-подзолистыми и торфянисто-подзолистыми почвами.
В Саянах и на Алтае в поймах рек предгорных степей господствует тополь лавролистный, отличающийся высокой зимостойкостью. Далее с высоты 350-400 м над уровнем моря степь сменяется березовыми, осиновыми лесами с участием пихты. До высоты 800-900 м встречаются осиновокедровые леса. Над ними на высоте до 1300-1500 м над уровнем моря во влажных (осадков 500-1600 мм в год) и относительно теплых горных районах распространены горные черневые темнохвойные леса. Выше до 2300 м над уровнем моря проходит пояс высокогорных подгольцевых пихтовокедровых, лиственнично-кедровых лесов.
Классификация лесов может производиться в соответствии с лесорастительным районированием, которое используется для разработки региональных систем лесохозяйственных мероприятий.
Необходимость лесохозяйственного районирования для условий России была доказана (в начале 20 века) Г.Ф. Морозовым и Г.Н. Высоцким, В.Н. Сукачевым и др. исследователями.
В лесорастительном районировании (с 1977 г) используют следующие таксоны: для равнинной территории – область, провинция, округ. При этом область – азональный регион, зона и подзона – территория с границами широтного простирания, провинция – территория с границами меридионального простирания, каждая провинция подразделяется на округа по изменению примеси сопутствующих пород. В пределах округа предполагается выделять районы по группам коренных типов леса или их сочетаниям.
Лесохозяйственное районирование может не совпадать с лесорастительным так как оно должно отражать и однородный уровень лесного хозяйства, то есть основываться на лесоэкономическом районировании. А.А. Цымек разделил территорию страны по экономическим условиям на 4 зоны: леса промышленного значения, промышленно-защитные леса, зона малолесных районов и безлесная зона. В пределах их он выделил 26 подзон, границы которых не совпадают с лесорастительным районирование.
При лесохозяйственном районировании границы согласуются с лесорастительными делениями. По Б.П. Колесникову, это разделение территории на единицы с однородными способами ведения лесного хозяйства и лесоэксплуатации.
1.Как влияет климат на распространение лесов в нашей стране, и каковы закономерности влияния климатических факторов на производительность древостоев?
2.В чем заключается разнообразие лесов на земном шаре?
3.Расскажите об основных лесорастительных подзонах нашей страны.
4.Какие закономерности наблюдаются в вертикальной зональности лесов?
5.В чем вы видите суть леса как явления географического и исторического?
Основная литература:
Мелехов И.С. Лесоведение. М.: МГУЛ.- 2004, 398 с.
Дополнительная литература:
Погребняк П.С. Общее лесоводство. Колос. М.: 1968, 440 с.
1Тепло и свет как экологические факторы
2Свет и продуктивность древостоев
3Распределение света в лесу
4Отрицательное действие низких и высоких температур
Основным источником тепла является солнечная радиация. Кроме того, тепло в приземную атмосферу поступает из глубинных слоев земли в результате распада радиоактивных веществ, разложения органических остатков, а также других источников.
На разных этапах развития (начало сокодвижения, распускание почек, прорастание семян, цветение, образование завязи, созревание плодов, начало фотосинтеза, рост), древесным породам необходимы определенные тепловые условия.
Ассимиляция лесных растений происходит при температуре от 0 до 40500С, достигая максимума при 25-300. У разных пород крайние и средние точки ассимиляции различны, но у большинства они находятся в указанных пределах. Дыхание древесных пород происходит и при температуре ниже 00, хотя оптимальной является температура 45-500, при 550 дыхание прекращается.
У многих растений семена начинают прорастать при температуре воздуха чуть выше 00, стебель и листья образуются при 60, а цветение при температуре выше 150. С повышением температуры воздуха и почвы становится более интенсивной деятельность микрофлоры, что способствует лучшему усвоению древесной растительностью азота, фосфора, и других веществ.
Тепло как экологический фактор играет огромную роль в жизни леса. При этом большое значение играет микроклимат, особенно в приземном слое воздуха и почвы. В этом слое теплообмен, передача тепла происходит в 4-х различных формах: молекулярная теплопроводность, обмен или турбулентная диффузия, тепловое излучение, передача тепла водой в ее различные состояния.
Свет – необходимый фактор жизни растений. При участии света в листьях образуется хлорофилл, происходят фотосинтез, транспирация, а также формирование и рост почек, тканей, листьев, цветков, плодов, обмен веществ между клетками и т. д.
Солнечный свет в лесу влияет на листообразование, ветвление кроны, ее форму и размеры, на форму ствола и, очищение от сучьев, интенсивность естественного изреживания древостоя, разложение подстилки, величину прироста и качество древесины, а также на плодоношение деревьев и урожай семян.
Преобладающая часть лесных растений нуждается в свете. Потребность в свете – генетически закрепленный комплекс структурных признаков и функциональных особенностей организации вида. Взаимоотношения растений со светом управляются генетически, вместе с развитием растений меняются его потребности в световой энергии.
Лесовод – ботаник И. Визнер (1907) создал классификацию типов освещения в условиях лесного биогеоценоза. Он различал следующие виды освещения:
1. Верхнее – падающее сверху на крону и листья растений.
2. Боковое – падающее косо или горизонтально на вертикальную площадь. Источником бокового освещения может быть Солнце, небо или отраженный свет (от деревьев, стен, и т.п. от 1:2 до 1:4).
3. Сквозное – представляющее собой лучи, проникающие внутрь леса сверху через кроны, листья, просветы между ними.
4. Нижнее – отраженное от поверхности почвы или воды.
Дневной свет слагается из прямого солнечного света и рассеянного, отраженного небосводом. Для фотосинтеза большое значение имеет рассеянный свет.
Различные лучи Солнца служат разным целям: красные, оранжевые и желтые используются растением для усвоения углекислого газа и образования хлорофилла, фиолетовые, синие и голубые для роста и развития почек. Транспирация осуществляется под влиянием всех лучей, но преимущественно желтых.
Потребность растений в свете всегда выше зимой, чем летом. На этом основании еще в начале прошлого столетия составлялись экологические шкалы отношения древесных пород к свету.
Наиболее старым и вместе с тем надежным является метод определения светолюбия древесных пород по внешним признакам.
Наряду с визуальными методами определения светолюбия древесных пород существуют специальные методы (или объективные), которые основываются экспериментальными данными, связанными с применением аппаратуры, измерениями и вычислениями.
К специальным методам относятся:
1) фотометрические, связанные с непосредственными замерами освещенности. Один из таких методов предложен И. Визнером (1907), предложен на различии в потемнении фотобумаги, помещенной в нижней части крон разных древесных пород.
Этот метод широко применялся в Австрии, Германии, Швеции для установления минимума светового довольствия древесных и недревесных растений.
Недостаток метода: при нем улавливаются преимущественно синий и фиолетовые лучи, а для фотосинтеза важной является желтая и красная часть спектра.
Л.А. Иванов изобрел специальный прибор фитоактинометр, который учитывает активные в фотосинтезе лучи. В настоящее время излучение при фотосинтезе (в пределах длины волн от 380 до 700 нм) принято называть фотосинтетически активной радиацией (ФАР).
2)физиологические методы, связанные с изучением физиологических процессов. К ним относится метод В.Н. Любименко, который заключается в том, что изучается чувствительность к свету хлорофильного аппарата различных древесных пород и таким образом определяется степень их теневыносливости. Его опыты доказали физиологическую обоснованность лесоводственного разделения древесных пород по их отношению к свету.
Л.А. Иванов и Н.Л. Коссович (1932) изучали фотосинтез у разных древесных пород методом ассимиляционных колб, позволяющим исследовать интенсивность фотосинтеза листьев и ветвей или всей надземной части небольшого растения без нарушения их естественного положения.
3)фитометрические методы, связанные с замерами самих деревьев. Метод Я.С. Медведева или таксационный метод относительной
высоты.
Метод М.К. Турского и В. Никольского основан на опытах с затенением сеянцев сосны и ели на грядах. Путем разной дозировки освещенности опытным путем подтвердили большее светолюбие сосны по сравнению с елью.
Опыт Цизляра (1904) австрийского исследователя применившего вышеуказанный метод с большим количеством пород и вариантов показал, что умеренное затенение может положительно сказаться на теневыносливой породе, и, наоборот, любое затенении сказывается отрицательно на породах более требовательных к свету.
4) анатомические методы, связанные с исследованием анатомического строения частей исследуемых деревьев.
Русский лесовод И. Сурож (1891) проводил измерения палисадной и губчатой паренхимы листьев у большого количества видов древесных и кустарниковых растений. По результатам этих исследований он составил классификацию светолюбия и теневыносливости древесных пород в виде определенного ряда – от наиболее теневыносливых (тис) до наиболее светолюбивых (сосна горная).
В современных условиях анатомические методы приобретают большое значение.
М.К. Турский на материалах наблюдений и обобщения опыта многих исследователей предложил шкалу светолюбия древесных пород. В этой шкале он расположил древесные породы по мере уменьшения светолюбия: лиственница, береза, сосна обыкновенная, осина, ива, дуб, ясень, клен, ольха серая, ильмовые, сосна крымская, ольха черная, липа, граб, ель, бук, пихта.
От света во многом зависит внешний облик деревьев. Форма кроны и ствола, количество побегов, их появление, рост и отмирание связаны с условиями освещения. Большая часть органического вещества растения создается в процессе фотосинтеза.
В общем виде связь между фотосинтезом и продуктивностью выражена Л.А. Ивановым (1976) и выглядит так:
М + м = IP T – a P1 T1
где: М – масса растений; м – вес отмерших частей;
I – интенсивность фотосинтеза; P – рабочая поверхность листьев; T – время работы фотосинтеза;
a – интенсивность дыхания; P1- масса живых частей;
T1 – время работы дыхания.
Максимальная продуктивность фотосинтеза свойственная зимне-голым лиственным породам, находится между 5 и 10 СО2 в час на 1 дм2 удвоенной площади листа. У вечнозеленых деревьев максимальная интенсивность фотосинтеза достигает 4-9 мг, у хвойных 2-4 мг. В среднем в лесах умеренного пояса фотосинтез при лучших условиях погоды составляет не более половины упомянутых показателей.
Свет – фактор, наиболее поддающийся регулированию со стороны лесовода. Можно изреживать густой лес для увеличения прироста в толщину отдельных деревьев или для появления подроста под пологом старого леса; создавать окна в пологе, чтобы получать групповой подрост разного породного состава.
Свет, влияя на развитие крон деревьев, воздействует на их плодоношение. Деревья хорошо освещенные, с хорошо развитыми кронами плодоносят лучше затененных со слабо развитыми кронами, из которых деревья пятого класса по Крафту совсем не дают семян, 4 класс – очень малое количество.
В пределах одного дерева плодоношение в разных частях кроны не одинаково. Плодоносят обычно освещенные части дерева.
Восточная и юго-восточная части кроны ели дают большее количество семян и лучшего качества.
Следует учитывать и географическое расположение. В таежных лесах в отличие от Воронежской лесостепи наибольшее количество шишек, как правило, образуется в верхней части кроны.
Свет, попадая в сферу влияния леса, изменяется количественно и качественно. Часть световых лучей отражается от полога, часть проходит через просветы между кронами, листвой и хвоей в пределах кроны, часть света проникает сквозь листву. Лучи достигшие поверхности почвы, часто поглощаются ею, частью отражаются от нее.
Световая обстановка в лесу определяется преимущественно древостоем, его составом, возрастом и высотой, сомкнутостью, густотой, ярусностью, размещением деревьев. Особенно большое значение имеет характер крон, их глубина, ширина, ажурность, плотность, размещение; почвенно-климатические условия и т. п. Количество света под пологом обусловлено временем года, особенно для лиственных пород.
В высокосомкнутых еловых насаждениях к поверхности почвы света попадает до 5-10% полного освещения. Спелые сомкнутые одноярусные древостои в период вегетации пропускают сквозь полог не более 40-50% световых лучей. При наличии второго яруса древостоя количество света, доходящее до поверхности почвы, резко падает.
Уменьшение полноты и сомкнутости полога, частичное или полное удаление подлеска, воздействие на другие ярусы приводит к увеличению количества света в лесу.
Состав света, проникающего под полог леса, обеднен физиологически активными лучами по сравнению с их составом на открытом месте.
Древостои из теневыносливых пород пропускают меньше света, чем древостои из светолюбивых пород.
Лесоводственные методы регулирования освещенности в лесу
Многие лесохозяйственные мероприятия направлены на регулирование освещенности в лесу и на создание в нем светового режима, соответствующего требованиям древесных растений в разные периоды их жизни. Можно изреживать густой лес для увеличения прироста отдельных деревьев в толщину или для появления подроста под пологом старого леса; создавать окна в пологе леса, чтобы получить групповой подрост разного породного состава; рубить спелый лес сплошными лесосеками и использовать полную радиацию для улучшения роста самосева и подроста.
Для лучшего роста молодняков в высоту и образования прямых и полнодревесных стволов необходимо боковое затенение.
В молодых и средневозрастных древостоях рубками ухода можно увеличить не только количество света, поступающее к оставшейся части деревьев, но и количество влаги и питательных веществ, что в свою очередь вызывает увеличение прироста.
К лесоводственным методам регулирования освещенности в лесу относится регулирование состава пород.
Понятие низкие температуры не всегда означают абсолютное выражение температуры – большой абсолютный минимум. Надо учитывать: конкретные условия среды (климатические, почвенные и пр.); природу данного растения, его предшествующую историю; рост и развитие; возраст растения, ход изменения температурных условий во времени, особенно до и после минимального значения температуры воздуха.
Рассмотрим отрицательное действие низких температур последовательно по мере его появления в течение года.
Поздневесенние заморозки возникают обычно в начале вегетационного периода. Чувствительны к ним всходы, молодая хвоя, листья и новые побеги некоторых видов. Клетки погибают или от внутриклеточных кристаллов льда, или от обезвоживания при образовании льда в межклетниках и оболочке клеток.
Рано распускающиеся листья и молодые побеги бука, клена, ясеня, ильма гибнут даже при слабых заморозках, когда температура воздуха снижается до – 0,50 – 2,00С. Следует иметь в виду, что при радиационных заморозках воздух охлаждается от листьев и побегов, которые имеют температуру ниже, окружающего воздуха.
При медленном понижении температуры древесные растения проходят один из видов морозной закалки, во время которой вода успевает выйти из клеток и лед образуется только в межклетниках.
Поздней осенью и при оттепелях зимой молодые растения гибнут от
выжимания корней кристаллами льда, которые образуются в почве до глубины 10 см поднимая ее. При оттаивании почва опускается, а корни оказываются на поверхности, и не могут удержать в вертикальном положении самосев или сеянец, который погибает. Иногда кристаллы льда выходят на поверхность, увлекая за собой корни всходов и все молодое растение.
Зимой гибель деревьев может произойти от сильных морозов. При этом более чувствительной оказывается корневая система, поскольку она продолжает расти даже глубокой осенью. Корневая система ели европейской погибает при температуре – 330С, а надземная часть ее выносит морозы до –520С.
Чередование морозных дней с оттепелями усиливает вред от ранних заморозков.
Частичное обмерзание крон лиственных пород (дуба, ясеня, клена и др.) может произойти не только из-за сильных морозов, но и вследствие зимней засухи, возникающей под влиянием сильных ветров и частой смены морозов оттепелями.
Морозобойные трещины появляются при резком снижении температуры и термическом сжатии периферийной зоны ствола.
При их образовании слышен треск деревьев, потому, что внутренние части древесины вследствие ее плохой теплопроводности сохраняют прежний объем.
Часто трещины образуются у дуба, ясеня, липы, граба, осины, береста, клена полевого и др. Мелкие трещины зарастают, но могут в следующие зимы еще более расширяться от морозобоя, вызывая снижение товарной ценности древесины, появление грибных инфекций и ослабление дерева, что способствует нападению вредных насекомых.
В конце зимы или в марте под воздействием прямой солнечной радиации и ночных морозов в нижней, наиболее темной части ствола в стене леса и у волока возникает солнечно-морозный припек ствола. При этом клетки флоэмы, камбия погибают днем от набухания, а ночью от замерзания в них воды, либо оболочки клеток разрываются из-за быстрого возвращения воды в межклетники при образовании там кристаллов льда. Затем развивается сухобокость ствола. Она расположена с южной или югозападной его стороны и начинается выше уровня снега. Солнечный припек встречается на стволах бука, дуба, ясеня, липы, осины и др. пород.
Ранней весной наблюдается солнечно-морозный припек хвои. Под влиянием прямой солнечной радиации хвоя оттаивает, ткани активизируются, а ночью мороз вызывает замерзание воды и гибель клеток. Солнечно-морозный припек хвои встречается у опушечных деревьев сосны обыкновенной, ели, но больше всего страдает пихта.
Поврежденные в результате неблагоприятных метеоусловий ткани, отдельные органы и молодые растения поражаются инфекционными болезнями, которые ускоряют их гибель.
Древесные растения наиболее чувствительны к высоким температурам в молодом возрасте. Всходы ели погибают при температуре 53-560С.
Температура, при которой погибает камбий всходов и самосева сосны, выше на 5-60С.
Летом под воздействием высоких температур может произойти опал корневой шейки. Это самый распространенный случай гибели древесных растений по этой причине. Опал наблюдается на открытом месте в жаркий день, когда почва сильно нагревается, и от нее температура тканей не одревесневшего стебля самосева и сеянцев повышается до + 500С и выше. Ожог вызывает денатурацию белков, наступает самоотравление токсическими продуктами обмена, приводящие к гибели клетки. Ткани корневой шейки в месте опала краснеют, буреют, образуется перетяжка, растение падает и погибает.
Опал усиливается не только на почве темного цвета, но и на песках. Поэтому мульчирование посевов песком недопустимо. Меры предосторожности: применение опилок, торфа, щитов в питомниках и разбрасывание порубочных остатков на вырубке.
Подобные ожоги, но только от прямого воздействия солнечных лучей, называется солнечными ожогами коры. Они наблюдаются у освещенных с южной и западной стороны тонкокорых и темноцветных стволов ели, бука, пихты, дуба, граба, ясеня, явора.
Ожог хвои (листьев) наблюдается у ели, пихты, клена, каштана. От высоких температур в питомниках могут поражаться листья бука, дуба, липы, а также сеянцы ели, лиственницы.
1. Какие вы знаете методы определения светолюбия?
2. Как распределяется в лесу свет и тепло?
3. Какое значение имеют циклы солнечной активности в жизни леса?
4.Как влияют высокие и низкие температуры воздуха на древесные и кустарниковые растения?
5. Какова взаимосвязь между светом и продуктивностью древостоя?
Основная литература:
Мелехов И.С. Лесоведение. М.: МГУЛ.- 2004, 398 с.
Дополнительная литература:
Погребняк П.С. Общее лесоводство. Колос. М.: 1968, 440 с.
1Материально-энергетический обмен
2Влияние леса на газовый состав атмосферы
3Особенности лесного воздуха
4Влияние загрязнения атмосферы на лес
5Лес и ветер
6Лес и влага
7Отрицательные явления, связанные с влагой
8Отношение древесных пород к влаге
9Водный баланс в лесу
Взаимодействие атмосферного воздуха и леса проявляется в основном в круговороте углерода и кислорода через процессы фотосинтеза и минерализации органических соединений, в действии атмосферных загрязнений на лес и взаимном влиянии ветра и леса.
Атмосферный кислород используется живыми организмами в лесу для дыхания и пополняется в результате фотосинтеза.
Глобальную проблему представляет накопление в атмосфере углекислого газа. Стволовая древесина наполовину состоит из углерода, значительную часть составляет кислород. Синтез древесины (природного полимера) происходит в результате материально-энергетического обмена лесного биогеоценоза с внешней средой
На образование 1 т абсолютно сухой стволовой древесины требуется 1,84 т. СО2, 0,54 т. Н2О и 0,01 т. минеральных веществ, т. е. 2,39 т. вещества.
Разница между этой массой и 1 т. полученной древесины составляет кислород (1,39 т), выделяемый в атмосферу в процессе фотосинтеза. При этом не учтена масса воды, участвующая в фотосинтезе, дающей кислород и составляющей незначительную часть от всей транспирируемой влаги.
Количество транспирируемой влаги у разных древесных пород различно и зависит от температуры, влажности воздуха, запасов влаги в почве и других факторов. По расчетам С.Б. Белова на транспирацию расходуется 11·108 кДж солнечной энергии, которая представляет собой скрытую теплоту парообразования и выделяется при конденсации паров и выпадении осадков. На фотосинтез, при котором энергия квантов солнечного света (фотонов) переходит в энергию химических связей органического вещества, затрачивается 21·106 кДж, и эта энергия выделяется при минерализации древесины, например при ее сгорании, во время пожара. Эта схема материально-энергетического обмена не учитывает в полной мере все его затраты.
Общей фитомассы создается в год на 1 м2 в бореальных лесах 0,65 кг, в смешанных лесах умеренного теплого пояса 1 кг, в тропических дождевых лесах – 2,8 кг.
Луга умеренных широт дают 0,8 кг/м2, культивируемые земли 0,65 кг на м2и болота 2,0 кг/м2.
Из этого следует, что годовой материально-энергетический обмен лесов выше, чем у луговых сообществ и некоторых сельскохозяйственных культур и ниже материально-энергетического обмена большинства болот. В лесу, в отличие от болот не происходит накапливания мертвых органических остатков из-за высокой кислородопродуктивности.
Сухой атмосферный воздух у поверхности земли содержит по объему 78% азота, 21% кислорода, около 1% благородных газов (аргон, неон и др.), 0,033% СО2, 0,01 % водорода.
Такого количества СО2 недостаточно для максимальной продуктивности древесных растений, тем более, что на высоте крон его содержание вследствие потребления в процессе фотосинтеза уменьшается до
0,02 %.
В надпочвенном слое лесного воздуха особенно ночью в сырую погоду концентрация СО2 доходит 0,07-0,18 % так как он выделяется в результате
дыхания корней, почвенной фауны, микроорганизмов и других процессов. Особенно много двуокиси углерода в начале вегетационного периода из-за обилия прошлогоднего опада и достаточного тепла и влаги для его минерализации. Повышенное содержание СО2 улучшает жизнедеятельность подроста, что компенсирует недостаток света.
Концентрация СО2 в лесу уменьшается в связи с неравномерностью его потребления в суточном и годичном циклах, а также в зависимости от высоты над поверхностью почвы. Максимальное содержание СО2 в области крон наблюдается летом. Осенью оно понижается. Максимальная концентрация СО2 приходится на припочвенный слой воздуха, минимальная
– на полог крон деревьев. В приземном слое воздуха концентрация СО2 зависит от породы, типа леса, возраста, полноты, сомкнутости древостоя, скорости ветра, рельефа местности, влажности почвы.
Наибольшее количество СО2 в лесу поступает в воздух в результате минерализации лесной подстилки; разложения органических веществ, гумусового горизонта почвы; жизнедеятельности микроорганизмов и грибов; дыхания растений, животных и человека (человек за сутки выделяет 1,2 % СО2 от стволовой массы) притока из атмосферы.
Кислород необходим для дыхания. Убыль атмосферного кислорода вследствие дыхания, гниения и горения возмещается кислородом, выделяющимся при фотосинтезе.
Леса России ежегодно выделяют более 4 млд. Тонн кислорода, поглощая при этом около 5,5 млд т. СО2. В частности 1 га сосновых лесов выделяет ежегодно 30 т., лиственных – 16 т., сельхзкультуры – 3-10 тонн кислорода.
В составе лесного воздуха кислорода не больше, чем в поле или на вырубке, но он более ионизирован. На поверхности земли ионизация молекул газов происходит под воздействием веществ в почве и электрических зарядов атмосферы и земли.
Растительность, особенно лесная, усиливает их взаимодействие. Днем во время вегетационного периода ионизация выделенных молекул кислорода происходит в основном за счет фотоэлектрического эффекта на поверхности листьев при фотосинтезе. Утром и вечером ионизация усиливается за счет газообразной фракции продуктов полураспада радиоактивных веществ, находящихся в земле.
Лес ионизирует кислород, выделяемый при фотосинтезе и приносимый ветром.
В лесу в результате окисления смолистых веществ образуется озон, а сами смолистые вещества относятся к фитонцидам. Открыл фитонциды в в 30-х годах 20 века советский биолог Б.П. Токин. По его определению фитонцидами называются продуцируемые растениями бактерицидные, фунгицидные и протистоцидные вещества, являющиеся одним из факторов их иммунитета и играющие роль во взаимоотношениях организмов в биогеоценозах. Иммунное значение фитонцидов проявляется при ранении живых тканей растений, в которых белки и углеводы были бы съедены микроорганизмами, но выделяемые фитонциды убивают их.
К фитонцидам относятся алкалоиды, глюкозиды, дубильные вещества, эфирные масла и другие соединения, химическая природа которых мало изучена.
Бактерицидное действие летучих фитонцидов различных видов растений определяется по скорости умерщвления микрооорганизмов.
По степени фитонцидности все древесные и кустарниковые породы подразделяются на:
1)Очень сильно фитонцидные – дуб черешчатый, клен остролистный.
2)Сильно фитонцидные – береза повислая и пушистая, сосна обыкновенная, ель европейская, осина, лещина, черемуха обыкновенная, можжевельник.
3)Средне фитонцидные - лиственница сибирская, ясень обыкновенный, липа мелколистная, ольха черная, кедр сибирский, рябина, акация желтая, сирень обыкновенная, жимолость татарская.
4)Слабо фитонцидные – вяз, бересклет бородавчатый.
5)Наименее фитонцидные – бузина красная, крушина слабительная.
Из кустарниковых и травянистых растений высокими фитонцидными свойствами обладают (в порядке убывания) багульник, чеснок, лук, горчица, крапива двудомная, ландыш.
Кроме летучих фитонцидов, лес является интенсивным производителем твердых примесей (аэрозолей) в виде пыльцы. Ежегодно 1 га леса может выбрасывать в атмосферу от 60 до 700 кг пыльцы, которая не обладает аллергическими свойствами.
Лес – незаменимый пылевой фильтр. В жаркие дни над его территорией наблюдается нисходящий поток воздуха, пыль оседает на кронах, затем дожди смывают ее в почву. 1 га леса может отфильтровывать из воздуха до 50-70 т. пыли в год.
Лес повреждается в районах заводов, фабрик, рудников, электростанций, работающих на буром угле от сернистого газа, серного ангидрида, фтористого водорода, фтора, хлористого водорода, сероводорода, аммиака и других соединений.
Наиболее вредными для древесной растительности и человека является серный и сернистый ангидрид, фтороводород, сероводород, аммиак и ацетилен. Воздействие сернистого ангидрида и фтороводорода проявляется в виде ожогов и последующего отмирания хвои и листьев. Токсичность фтороводорода в 10 раз выше токсичности сернистого ангидрида. Накопление окислов меди, цинка и свинца вызывает отмирание хвои и ее опад. Наличие в воздухе цементной пыли с высоким содержанием извести и кремневой кислоты вызывает закупорку устьиц и при проникновении щелочных элементов в ткани листа происходит подщелачивание клеточного сока листьев, разъедание покровных тканей.
По степени газоустойчивости все древесные и кустарниковые породы подразделяются на 3 категории:
1.Очень устойчивые – ель колючая, туя западная, робиния лжеакация и др.
2.Средней устойчивости – можжевельник обыкновенный и виргинский, лиственница сибирская, бархат амурский, береза пушистая и бородавчатая, дуб черешчатый и красный и др.
3.Неустойчивые – ель, пихта, сосна, каштан конский, рябина, ясень.
Особо следует остановиться на отношении леса к радионуклидам, так как лесные биогеоценозы являются одним из наиболее радиочувствительных типов природных комплексов. Самым неустойчивым к действию ионизирующей радиации является зооценоз, затем древостой (особенно хвойные породы) наиболее устойчивы мхи, лишайники.
Низкие дозы радиоактивности могут привести к образованию вторичных побегов, то есть деревья реагируют на них как на слабые яды, стимулированием роста. У млекопитающих радиация в 5-10 раз ускоряет развитие, повышает численность потомства. Но это надо рассматривать как патологическое явление.
При повышении дозы радиации и срока ее действия у растений отмечается аберрация хромосом, замедляется вегетативное развитие.
В первое время после радиоактивного поражения лес как фильтр задерживает на кронах 40-90 % радиоактивной пыли и общая радиоактивность под его пологом в десятки раз ниже, чем на открытом месте. Постепенно радиоактивные примеси попадают в подстилку, проникают через кутикулу листьев и включаются в обмен веществ растений.
Аккумулируя радионуклиды в коре, ветвях, древесине, лес препятствует их разносу по земной поверхности. Но с этого времени под пологом леса радиационное заражение становится выше, чем на открытом месте. Самоочищение происходит медленно. Среди японских деревьев сохранившихся вблизи ядерного взрыва, еще и сейчас оставляют на фотобумаге ряд траурных колец.
Ветер способствует опылению цветков и распространению семян большинства древесных пород лесной зоны, усиливает приток воздуха, обогащенного углекислым газом и при скорости до 2-3 м/с повышает фотосинтез, регулирует транспирацию.
Умеренный ветер при скорости больше 5 м/с усиливает испарение растением влаги и отрицательно влияет на производительность леса.
При одностороннем действие ветра формируется однобокая крона, так как с наветренной стороны молодые побеги усыхают. Этому могут способствовать и снежные бури. Если усыхают верхушечные почки, то рост ствола в высоту прекращается, и деревья начинают ветвиться. Так возникает стелющаяся форма деревьев – криволесье.
Ветер вызывает кривизну стволов, при раскачивании в них могут возникнуть трещины, произойти обрывы корней и охлестывание ветвями вершин ниже растущих деревьев.
При скорости ветра выше 10 м/с возникают черные бури. Ветры силой в 6 баллов (10-12 м/с) и выше причиняют лесу огромный вред, вызывая ветровалы и буреломы на больших площадях.
От ветровала и бурелома в древостое, не тронутом рубкой, больше страдают самые крупные деревья, а в разреженных насаждениях – средние и мелкие.
На ветровальных местах поверхность почвы неровная. Такие изменения рельефа, не превышающие нескольких дм, называются нанорельефом (по старой терминологии – микрорельефом).
Поэтому наличие нанорельефа в лесу указывает на прошлые ветровалы и опасность новых.
Повышенной ветроустойчивостью обладают смешанные насаждения. Ель – неустойчива против ветра из-за поверхностной корневой системы. Устойчивость придает ельникам примесь сосны объемом в 1/3 состава древостоя.
Ветер влияет на форму поперечного сечения ствола: из круглой она становится овальной. При этом годичные слои (креневая древесина) образуется у хвойных со стороны крена, а у лиственных пород – на наветренной стороне (тяговая древесина).
Лес в свою очередь также влияет на ветер. При движении в сторону леса скорость ветра на расстоянии около 60 м от опушки ослабевает примерно на 20-60 %, а также наблюдается сильный ток воздуха вверх. Глубина внедрения ветрового потока в лес во многом зависит от ветроупорной опушки, повышающей устойчивость ветровальных пород
(ель).
Ветроупорная опушка – полоса леса, предназначенная для защиты леса от ветровала. Закладывается преимущественно из лиственных древесных пород с глубокой корневой системой, способных развивать мощную крону.
Врываясь в насаждение, ветер постепенно теряет свою силу, расходуя ее на трение воздуха о стволы и ветви, а также на раскачивание стволов,
ветвей, листьев. Затухание скорости ветра в лесу зависит от древесной породы, полноты, высоты, формы древостоя, степени облиствения деревьев. Максимальная скорость ветра в лесу наблюдается над кронами деревьев, ближе к кронам она уменьшается, внутри крон затухает, а у поверхности почвы равна нулю.
За лесным массивом скорость ветра восстанавливается до первоначальной на расстоянии от 5 до 15 кратной высоты древостоя.
Источниками влаги для растений являются:
1. Атмосферные осадки – дождь и снег.
2. Конденсационные осадки в виде росы, инея, ожеледи, образующиеся
в результате сгущения водяных паров, капелек тумана, дождя.
3. Почвенная влага, образующаяся от предыдущих источников.
4. Грунтовые воды.
5. Реки и пресные водоемы.
Вода составляет половину массы ствола дерева и большую часть живых тканей.
Для переноса минеральных веществ в ионном состоянии, поддержании клеточного тургора, процесса фотосинтеза, охлаждения поверхности листьев (хвои) 1 га леса расходует за вегетационный период более 1 тыс. литров воды. Вода по растению непрерывно движется по системе: почва – растение
– атмосфера. Но если водный потенциал почвы при ее высыхании возрастает и сравнивается с водным потенциалом корней, то влага в растение не поступает.
Вода к растениям поступает не только из почвы. Тургор клеток часто поддерживается внешней влагой (дождь, роса).
Жизнь леса тесно связана с повышенной влажностью воздуха (60-90%). В вегетационный период под пологом леса по вертикали наблюдается два минимума влажности воздуха. Первый отмечается у поверхности почвы и обусловлен поступлением водяного пара из почвы и при транспирации воды живым напочвенным покровом. Второй максимум характерен для средней части древесного полога и образуется за счет транспирации деревьев.
Высокая влажность воздуха является необходимым условием для произрастания многих лесообразующих пород (бук, пихта, ель, кедр и др.).
Прорастание семян на почве возможно при определенном уровне ее влажности, характерном для каждого вида древесной породы и даже экотипа.
Физиологическое значение влаги проявляется также в извлечении питательных и минеральных веществ из почвы, превращение и передвижение их происходят при участии воды; влага играет большую роль в превращении протоплазмы и т. п.
Отрицательными явлениями в лесу считают зимнюю, весеннюю и летнюю засуху, заболачивание, вымокание семян и всходов, затопление и подтопление леса, снеголом и снеговал, снежные лавины, ожеледь и град.
Зимняя засуха возникает при частых оттепелях, когда усиливается транспирация, а корневая система находится в мерзлом грунте и не может восполнить большие потери влаги в живых тканях.
Ранневесенние засухи специфичны для лесостепных и степных районов Сибири, где сильные ветры и резкие перепады температур, интенсивная инсоляция вызывает обесцвечивание хвои сосны взрослых деревьев и даже их усыхание. Этот вид засухи сочетается с солнечноморозным припеком хвои.
Летние засухи характеризуются обычно отмиранием отдельных деревьев, которое может усилиться в ельнике 4-5 лет за счет активизации стволовых вредителей и возбудителей корневых гнилей.
Косвенно засуха снижает устойчивость деревьев вследствие обильного семеношения, при котором расходуются не только запасы крахмала и соединения азота прошлых лет. Такова биологическая закономерность сохранения вида – увеличить потомство в критическое для жизни время.
В степи опадение листьев как реакция на засуху характерно для тополей.
Засуха снижает устойчивость деревьев к холоду и приводит к гибели, если последующая зима выдается морозной.
Чаще из древесных пород от засухи погибает ель европейская, сосновый подрост в спелых древостоях и другие хвойные и лиственные породы.
Распространенным явлением считается заболачивание почв. При интенсивном заболачивании происходит ослабление деревьев и заселение их стволовыми вредителями. При большом недостатке кислорода в почве у ряда древесных пород возникает вначале анаэробное дыхание, но вскоре наступает токсикоз от накопившихся не полностью окисленных продуктов обмена веществ, и дерево погибает.
Семена многих древесных пород теряют всхожесть после нахождения длительного времени в стоячей воде. Позже определенных сроков (у сосны через 40 дней, ели через 20 дней) наблюдается загнивание семян, которое в лесоводстве получило название вымокания семян. При полном покрытии почвы водой происходит вымокание всходов. У растений загнивают корни, а затем наступает гибель.
Во время вегетационного периода даже кратковременное летнее затопление лесов может вызвать необратимые явления у большинства древесных пород. Летнее затопление в течение 15-30 дней переносят ива белая и ломкая, осина, некоторые виды тополей, дуб черешчатый, платан; болезненно – береза, ольха, робиния лжеакация, и погибают даже при
кратковременном затоплении орех грецкий, клены ясенелистный и серебристый, сосна и др.
Подтоплением называют сохраняющийся во время вегетационного периода уровень почвенно-грунтовых вод выше 80-140 см с зоной аэрации соответственно меньше 20-80 см. При этом избыток воды приводит к нарушению дыхания корней, денитрификации, накопления метана и других токсических веществ и соединений в растениях. Возникает хлороз листьев и хвои: бледно-зеленый цвет переходит в желтовато-зеленый, а затем они желтеют и опадают.
Лес страдает также от твердых осадков. Мокрый снег, накапливаясь на ветвях промерзших деревьев, вызывает снеголом и снеговал.
При снеголоме обламываются крупные сучья и средняя часть стволов, а при снеговале стволы наклоняются или вываливаются с корнем.
Для ослабления снеговала и снеголома не следует допускать перегущения молодняков, а если это случилось, проводить осторожное прореживание.
В некоторых районах повторяющиеся ежегодно снежные бури, особенно с ледяной крупой, вызывают эрозию поверхности побегов и тонких стволов деревьев. Иногда крупный град ломает ветви и наносит раны верхней части стволов.
В южной части лесной зоны, в лесостепи при встрече влажного воздуха
с фронтом зимнего холода переохлажденные водяные пары образуют на промерзших ветвях слой прозрачного льда. Это явление называется ожеледь. Под тяжестью льда ветви и вершины обламываются. Иногда ломаются стволы, реже деревья выворачивает с корнем. Сильно повреждаются берест, ясень, робиния лжеакация. Устойчивыми к ожеледи являются абрикос, айлант, вишня, гледичия, груша, клены и др.
Слабые повреждения ветвей может вызвать изморозь, представляющая собой отложения на ветвях, хвое непрозрачного рыхлого льда.
В горных условиях древостои уничтожаются снежными лавинами, которые вываливают и обламывают стволы деревьев.
Хотя для большинства древесных пород оптимальной является 60-80% от полной влагоемкости почвы, некоторые виды хорошо растут и при полной насыщенности влагой и недостатке кислорода в почве, а другие – в условиях с малым количеством влаги. Первые называются гигрофитами, а вторые – ксерофитами.
К гигрофитам относится ива серая, ушастая, белая и ольха черная, способные расти на постоянно сырых почвах или при высоком уровне залегания грунтовых вод.
Ксерофиты обычно имеют приспособления для уменьшения транспирации: листья мелкие (саксаул, джузгун), белого цвета или с опушенной кожицей (лох, облепиха, ракитник русский), покрытые волосяным налетом (шелюга) и свойство сбрасывать их при засухе (акация желтая). Ксерофиты характеризуются развитой корневой системой, способностью быстро регулировать движение замыкающих клеток устьиц и хорошо удерживать воду, а также способностью цитоплазмы противостоять механическим повреждениям и свойствам белков свертываться лишь при высоких температурах.
Ксерофитами считаются сосна крымская и эльдарская, гледичия. Среднее положение по требовательности к влаге занимает
лиственница, сосна сибирская (кедр), пихта сибирская, осина, береза повислая, липа мелколистная, бук. Они называются мезофитами. Корни мезофитов в отличие от ксерофитов, обладают слабой водоудерживающей способностью. В засушливые годы они испаряют воду быстро и неравномерно.
Дуб черешчатый является представителем переходной группы – ксеромезофитов. В нее входят берест и другие виды.
Между гигрофитами и мезофитами находится промежуточная группа – гигромезофиты. К ним относятся ель, береза пушистая, ольха серая, ясень обыкновенный, ива козья, осокорь, смородина черная, черемуха. Гигрофиты быстро растут там, где их корни проникают в капиллярную кайму грунтовых вод.
Кроме требовательности как способности вида извлечь влагу при ее недостатке в почве, различают потребность древесных пород во влаге. Она характеризуется количеством транспирируемой влаги при образовании определенного количества сухого органического вещества.
Ряд убывания потребности во влаге: робиния лжеакация, береза, дуб, осина, ель, сосна. Одна и та же порода – робиния лжеакация – может расти на сухой почве как ксерофит и в то же время неэкономно расходовать влагу.
Приходная часть водного баланса в лесу может быть выражена формулой: З1 + О + П + С + К, где
З1 – запас воды в почве, надземной и подземной органической массе; О – осадки (вертикальные и горизонтальные); П – подземный приток; С – снег или сползшие снежные лавины;
К – конденсационная влага.
Расходуется влага в основном на транспирацию (30-60%). Где выпадает осадков более 700 мм, эта доля будет меньше.
Уровень транспирации связан с приростом древесины. Больше транспирируют средневозрастные древостои в период максимума прироста по запасу. Транспирация зависит и от состава пород. Интенсивность транспирации березы бородавчатой в 2,5 – 3 раза выше, чем у сосны.
Транспирация возрастает с увеличением влажности почвы. Максимум транспирации наблюдается на влажных почвах.
В величину транспирируемой влаги входят проникшие под полог и в почву осадки, а также запасы воды в почве, подземный приток, надземный приток от принесенного со стороны снега и конденсационная влага.
Главная часть осадков, задержанных кронами, испаряется, увлажняя атмосферу, особенно лесной воздух. Количество задержанных осадков зависит от древесной породы, сомкнутости крон, возраста древостоя, интенсивности и продолжительности выпадения осадков, силы ветра, температуры.
Задерживаемая кроной влага при ее испарении охлаждает листья и хвою, снижает дефицит влаги в воздухе. Мокрые листья ослабляют транспирацию.
Физическое испарение с поверхности почвы в лесу меньше, так как в летний период днем температура воздуха там ниже, чем на открытом месте, ветра нет, а относительная влажность выше. Снижает испарение и лесная подстилка.
Физическое испарение и транспирация составляют суммарное испарение. При общей закономерности превышения суммарного испарения лесом по сравнению с нелесной территорией на 10-30 % в лесостепной и степной зоне вне пойм суммарное испарение может быть ниже.
Расход воды на сток подразделяется на несколько частей: поверхностный сток, грунтовый сток и подземный отток, уносящие воду за пределы конкретного участка леса и сток в недра земли.
Поверхностный сток в лесу может полностью отсутствовать, но чаще он колеблется от 5 до 20% выпавших осадков, уменьшаясь в равнинных лесах к югу. По доле грунтового стока судят о водоохранной роли лесов.
Грунтовый сток и подземный боковой отток вместе с остальными каналами инфильтрации изменяются в лесу в больших пределах: от 2 до 55% уменьшаясь в направлении от тайги к южным европейским равнинным лесам.
Незначительная доля влаги расходуется при фотосинтезе и на увлажнение образовавшейся фитомассы.
Таким образом, формула полного водного баланса в лесу будет выглядеть следующим образом:
О + К + П1 + С1 + З1 = Т + Икр + И др + Ф + С2 + П2 + З2 Где приходная часть нам известна (см. выше), а в расходной:
Т- транспирация (20-60%)
Икр – испарение осадков, задерживаемое кронами (10-20%) Идр – испарение с почвы, напочвенного покрова (10-30%) С2 – сток поверхностный (0-30%)
П2 = почвенный отток (2-30%)
Ф – диссонация воды при фотосинтезе (меньше 1%)
З2 – запас воды на конец наблюдений в определенной толще почвы и в органической надземной и подземной массе;
Упрощенная формула водного баланса: О = Т + И + С + П.
Если учесть, что доля транспирации (Т) с испарением (И) в лесу колеблется от 40 до 80%, а на вырубке от 20 до 60%, то можно сделать вывод, что при нормальной влагообеспеченности лесные насаждения на суммарное испарение расходует влаги больше, чем травянистая растительность. Сельскохозяйственные культуры по своему суммарному испарению могут не отличаться от лесонасаждений. Различие в стоке сохраняется. На вырубках расход на поверхностный и грунтовый сток соответственно равны 10-40% и 5-20%, то есть по сравнению с лесом поверхностный сток возрастает.
1. Влияет ли лес на содержание кислорода в атмосфере?
2. Какие промышленные газы оказывают наиболее сильное влияние на насаждения и в чем оно выражается?
3. Укажите положительную и отрицательную роль ветра в жизни леса.
4. Чем отличается лесной воздух от городского?
5. Расскажите о значении влаги в жизни леса.
6. Как влияет лес на уровень грунтовых вод и сток?
Основная литература:
Мелехов И.С. Лесоведение. М.: МГУЛ.- 2004, 398 с.
Дополнительная литература:
Погребняк П.С. Общее лесоводство. Колос. М.: 1968, 440 с.
1 Значение почвы в жизни леса
2 Влияние почвы на корневую систему деревьев
3 Влияние рельефа почвы на компоненты леса
4 Биологический круговорот веществ
5 Роль леса в почвообразовании
6 Лес на многолетней мерзлоте
7 Потери плодородия почвы
В.В. Докучаев – основатель научного почвоведения указывал на 5 факторов почвообразования: климат, материнская горная порода, рельеф, растения и животные и возраст местности. Таким образом, лесные растения
участвуют в формировании почвы, повышают ее плодородие. Плодородие – свойство почвы обеспечивать растения зольными элементами, соединениями азота и водой для образования органических веществ. Оно зависит от температуры почвы и степени ее насыщенности кислородом для дыхания корней. При снижении его содержания рост древесных пород ухудшается. В целом в почве объем воздуха должен составлять не менее 10-15 %.
Из воды и углекислого газа растения получают водород, углерод и кислород, которые идут на построение древесины. Кроме них в состав основных продуктов жизнедеятельности растений входят азот, фосфор, калий, сера, магний и др. элементы.
Азот – основной химический элемент белков и аминокислот. Лесные почвы, как правило, бедны азотом в воде ионов нитратов или аммония, образующихся под воздействием микробиологических процессов. Интенсивность этих процессов зависит от температуры.
Фосфор – необходим растениям для нормального протекания фотосинтеза и дыхания как химический элемент, участвующий в превращении энергии. Фосфор повышает зимостойкость. Недостаточно фосфора обычно на кислых почвах, что вызывает задержку роста и сроков цветения растений.
Калий влияет на интенсивность ассимиляции углекислого газа, регулирует осмотическое давление в клетках, улучшает передвижение питательных веществ и воды в тканях, активизирует синтез сахаров, белковых веществ, усиливает устойчивость растений к холоду, засухе.
Производительность лесов зависит также от кальция. Он входит в состав клеточных стенок, влияет положительно на рост корней, улучшает структуру почвы, снижает ее кислотность и предотвращает вымывание необходимых для растений химических элементов.
Магний входит в молекулы хлорофилла, сера – в состав белков. Железо имеет значение для синтеза хлоропластов, поэтому при его недостатке листья светлеют. Для фотосинтеза необходим еще хлор и цинк. Всего растения потребляют около 60 химических элементов.
Ферменты и гормоны растений содержат микроэлементы. Недостаток бора на осушенных и удобренных торфянистых почвах приводит к гибели верхушечных почек и целых деревьев, отмиранию побегов и др. Недостаток марганца затрудняет образование витамина С. При недостатке меди и бора, древесные породы не плодоносят. Молибден защищает растения от токсического действия подвижного алюминия на кислых почвах, и входят в состав ферментов. Фтор улучшает условия питания растений, а йод – обмен веществ.
Лесная подстилка богата микроэлементами, и в лесу недостатка их нет. Но если лес создается на месте пашней, болота, то внесение микроэлементов ускоряет рост и повышает устойчивость деревьев.
Иногда избыток элементов питания может отрицательно влиять на лес. Например, при избытке азота образуется рыхлая древесина, а накопление
нитратов на осушенных низовых болотах может вызвать гибель от низких температур. Избыток влаги в почве приводит к особому формированию корневых систем деревьев.
Почве принадлежит основная роль в формировании корневой системы деревьев.
Важнейшие функции коневых систем заключается в следующем:
1.С помощью корней растения закрепляются в почве, получая тем самым механическую устойчивость.
2.Деревья развивают в почве мелкие и крупные корни. Мелкие корни поглощают влагу из почвы с растворенной в питательными веществами.
3.В корнях откладываются запасные питательные вещества. Они
– источник корневого давления, способствующего подаче воды в ствол и крону.
4.Корни принимают участие в обогащении почвы элементами питания.
5.Корни выделяют в почву (с помощью экзосмоса) большое
количество углеводов, органических веществ, играющих роль питательной среды для почвенных микроорганизмов.
По морфологическим признакам корни лесных деревьев делят на горизонтальные, вертикальные, и косовертикальные, стержневые и якорные, проводящие и сосущие.
На богатых почвах деревья имеют менее развитую корневую систему, так как из небольшого объема почвы можно извлечь необходимые элементы питания и воду. На бедных почвах отношение массы корневой системы к надземной части возрастает. Наибольшее количество корней находится в гумусовом горизонте, несколько меньше в иллювиальном.
Наиболее пластичной, меняющейся в зависимости от гранулометрического состава и влажности почвы, считают корневую систему сосны. В оптимальных почвенных условиях у сосны образуются мощные горизонтальные корни, развитый стержневой о более глубокие боковые корни. Основная масса корней находится в верхнем 20 сантиметровом слое почвы.
На сухих песчаных почвах с глубоким уровнем грунтовых вод сосна развивает поверхностную корневую систему.
На песках при глубине грунтовых вод 2-2,5 м она имеет развитый стержневой корень.
На заболоченных почвах сосна формирует поверхностную корневую систему.
Кроме влажности и биологических свойств пород на развитие корневой системы влияет плотность грунта. Отрицательно воздействуют на рост корней древесных пород легкорастворимые соли: сульфаты натрия и магния, нитраты кальция и магния.
Кроме того, на развитие корней влияют температура почвы, а также патогенные грибы, вредители корней, густота древостоя, деятельность человека и др. В зависимости от сочетания этих факторов корневые системы подразделяются на несколько типов. М.И. Калинин выделил 8 типов корневых систем, из которых наиболее выраженными являются 5.
Тип 1 – имеет стержневой корень, и нет вертикальных ответвлений от горизонтальных корней.
Тип 2 – отличается от 1 типа наличием вертикальных корней, не нарушая конусовидного габитуса всей системы.
Тип 3 – вертикальные корни достигают глубины стержневого корня, габитус корневой системы чашеобразный (например, на многолетней мерзлоте).
Тип 4 – доминируют якорные корни.
Тип 5 – стержневой корень отсутствует, якорные недоразвиты, корневая система образована в основном горизонтальными корнями (в очень сухих условиях лесной зоны).
Эту классификацию М.И. Калинин сопроводил цифровыми данными, рассчитав коэффициент ветвистости горизонтальных корней по формуле:
К = Σ I / L
где Σ I – суммарная длина скелетных (толще 3 см) ответвлений первых трех порядков, м
L – длина материнского горизонтального корня, м.
По данным М.И. Калинина, коэффициент ветвистости горизонтальных корней на Украине оказался у березы повислой 17,18, у ели европейской 5,65, у сосны обыкновенной 2,98, у липы крупнолистной 2,07, у клена остролистного 1,83, у дуба черешчатого 1,46.
Рельеф влияет на водный и тепловой режимы почвы. С ним связаны перераспределение выпавших атмосферных осадков, грунтовых вод, изменение в мощности и составе почвы, перемещении почвенных частиц и т. д. Все эти изменения накладывают от печаток на характер леса.
С.В. Зонн (1964) подразделяет почвы по их положению в рельефе и выраженности миграции веществ на следующие группы: 1. Элювиальную; 2. Транзитную и 3. надводно-подводную.
Почвы 1 группы формируются на водоразделах и характеризуются поступлением веществ и энергии только из атмосферы, глубоким стоянием грунтовых вод, недоступных для снабжения растительности. Почвы 2 группы приурочены к склонам с характерным для них перераспределением атмосферных осадков, световой и тепловой энергии, поступлением влаги и продуктов обмена веществ и энергии с прилегающих биогеоценозов, а также
дополнительной аккумуляции веществ из грунтовых вод. Почвы 3 группы лесных биогеоценозов приурочены к отрицательным формам рельефа, регулярно или периодически затопляемым водами. Для этих почв характерны: периодическое обогащение веществами с поверхности, выщелачивание подвижных элементов питания, торфянистость, иловатость и соленость.
Повышенным месторасположением свойственна элювиальная группа почв с их глубоким стоянием грунтовых вод и другими особенностями. Здесь формируются леса средней или низшей продуктивности, связанные с недостатком влаги и питательных элементов в почве.
Водный, тепловой и световой режимы склонов связаны с их экспозицией, что отражается и на составе лесов и их продуктивности.
Имеет значение и направление господствующих ветров. Наветренные западные склоны Урала получают больше осадков, и здесь распространены еловые и пихтовые леса, а на восточных склонах мало выпадает осадков, поэтому преобладают сосняки.
Высохшие, выпуклые формы рельефа находятся под более мощным воздействием сильных ветров, и объем ветровала и бурелома может быть больше именно здесь, а не в пониженных месторасположениях с избыточно увлажненными почвами, где у деревьев формируется поверхностная корневая система.
Лесовозобновление ослабляется с повышением крутизны склона. Даже нанорельеф влияет на возобновление под пологом леса.
Влияет на лесовозобновление и почва. Почва влияет как на производительность древостоев, так и на техническое качество древесины, а также на состав и форму древостоев. На богатых почвах формируются многоярусные смешанные древостои с развитым подлеском и травостоем, так как такие почвы являются оптимальными почти для всех лесных растений. При недостатке или избытке каких-либо веществ образуются чистые древостои.
На разных типах почв произрастают различные насаждения. Так на солонцеватых почвах лесостепи и степи чаще произрастают низкобонитетные дубравы. На карбонатных почвах поселяется лиственница, ясень и другие породы. На сухих известковых почвах может произрастать только сосна.
Под биологическим круговоротом веществ понимают поступление их из почвы и атмосферы в растение, биологический синтез с образованием сложных органических соединений, возвращение веществ из живых организмов в почву и атмосферу, и превращение их в доступные для растений соединения.
Влесу круговорот веществ наиболее мощный благодаря глубокому проникновению корней деревьев в почву и микотрофному питанию многих из них.
Первым звеном этого круговорота называют ризосферное звено. К ризосфере относится тонкий слой почвы вокруг корней, равный средней длине корневого волоска (3-4 мм). Иногда понимают под ризосферой всю корнедоступную глубину почвы. Ризосферная почва подкислена корневыми выделениями растений и очень богата микроорганизмами. Ризосферное звено
–это переход от мертвой природы к живой. Необходимо увеличивать объем ризосферной почвы путем создания сложных насаждений из древесных пород с различными типами корневых систем и поддержания глубокой аэрации почв (С-Е, С-Д, Лц-Е, Б-Е).
Вещества и химические элементы, поступающие через сосущие корни расходуются на образование органических веществ в процессе ассимиляции в листьях и хвое деревьев. Они также участвуют в синтезе сложных органических веществ. Следовательно, основное потребление минеральных веществ происходит в кроне, и там находится второе звено круговорота –
звено потребления.
Некоторые вещества вымываются из кроны и поступают в почву, но основная часть возвращается в виде органических остатков – опада и отпада.
Опавшие листья, хвоя, веточки, плоды, кора (1-7 т/га в год), отмершие корни (0,1 – 0,2 т/га) и стволы (без рубок 0,5-1,0 т/га) частично минерализуются под воздействием внешних условий. Основным разлагающим агентом являются кислоты, растворенные дождевой водой. При этом выделяется углекислый газ, выщелачивается калий, фосфор, в таежных лесах – кальций и магний.
Опавшие листья разлагаются под воздействием паразитных грибов (сапрофитов) и возбудителей болезней. Это звено первичного разрушения органического вещества.
Далее участвует почвенная фауна: дождевые черви, мокрицы, многоножки, клещи и др.
Вкишечнике и экскрементах дождевых червей образуются вещества, доступные для питания деревьев.
Под воздействием бактерий и актиномицетов сложные органические соединения превращаются в менее сложные. Все это звено называется звеном разложения и синтеза.
При вымывании продуктов разложения из подстилки в минеральную часть почвы появляются молекулы гумусовых соединений.
Гумус почвы является источником питательных элементов для растений благодаря своей высокой обменной поглотительной способности и возможности минерализоваться до простейших подвижных форм питания. Эта часть круговорота называется звеном гумификации и последующей минерализации.
Освободившиеся химические элементы, не вовлеченные в ризосферное звено, могут перейти в почвенный раствор и закрепиться в почве. Важным хранилищем питательных катионов являются коллоидные глины. Это последнее звено называется звеном необменного и обменного поглощения веществ почвой. Количество питательных веществ характеризует трофность (богатство) почвы и отражает емкость обмена или емкость поглощения почвы.
При биологическом круговороте веществ их поток проходит через растительноядных наземных животных и их хищников или через вредных насекомых. При разложении этих консументов вещества вновь поступают в лесную подстилку. Так завершается рассмотренный нами биологический круговорот веществ в лесу.
Иной характер разложения имеют отмершие корни, главным образом корневые волоски. Они отмирают постепенно по мере роста корня и образования новых волосков.
Таким образом, производительность леса зависит не только от валового содержания питательных элементов в почве, но и от интенсивности круговорота.
Лес, являясь аккумулятором веществ и энергии, играет большую роль в почвообразовании.
Биологический круговорот веществ проходит главным образом через верхние горизонты почвы, где накапливаются азотистые соединения и зольные вещества в доступных для растений форм. Поэтому у древесных пород, имеющих даже развитую стержневую систему (дуб, сосна) основная часть мелких корней расположена в гумусовом горизонте. На этом горизонте в первую очередь сказывается воздействие леса. Так, поселение дуба на дерново-подзолистой почве изменяет реакцию ее верхнего 10 см слоя в сторону нейтральной и увеличивает ее плодородие.
Благодаря глубоким корням деревья вовлекают в биологический круговорот все новые порции питательных веществ.
Высокими почвоулучшающими свойствами отличаются деревья и кустарники, на корнях которых распространены микробы-азотфиксаторы. Сюда относятся акация желтая, робиния, ольха серая, черная и др. Обогащение почвы азотом происходит в основном через опад. Кроме того, в процессе жизнедеятельности клубеньковых бактерий образуется аммиак, который растворяется в почвенных водах и может поглощаться растениями.
Положительная роль некоторых пород заключается в сильном разрыхлении почвы тонко разветвленной корневой системой. Листья ясеня, клена, граба содержат наибольшее количество зольных элементов. Опад дуба, бука, березы и др. пород имеет высокую концентрацию азота, а листья
липы, осины, ильмовых и бузины, обогащают почву кальцием и способствуют оструктуриванию.
Одним из факторов, влияющих на свойства почвы, является состав редуцентов. Основными разрушителями грубогумусной подстилки хвойных лесов средней и северной тайги являются пенициллиновые и мукоровые грибы, в результате жизнедеятельности которых образуются фульвокислоты.
С влиянием леса тесно связан подзолообразовательный процесс, особенно выраженный в таежной зоне – в лесах с преобладанием мохового напочвенного покрова, в условиях затрудненного разложения опада и подстилки, образования в повышенных количествах фульвокислот.
Длительное существование одной и той же хвойной породы на одном и том же месте усиливает процесс оподзоливания настолько, что он приводит к резкому падению продуктивности древостоев.
Не отмечен подзолообразовательный процесс и при большем количестве выпадающих осадков, но особенно в теплых условиях Центральной Европы.
Влияние леса на почву неравномерно, а в пределах одного биогеоценоза существует неоднородность местообитаний. Кислотность почвы повышается в направлении от периферии проекции кроны к основанию ствола, так как атмосферные осадки вымывают из кроны органические и минеральные вещества кислой природы. Большое значение при этом имеет опад коры, содержащий 3-4 % зольных элементов. Например, масса опавших чешуек коры в сосняке выше опада ветвей на 20-30 %.
Многолетняя мерзлота почвы представляет собой или остаточное явление оледенения суши, или возникает как предшественник нового ледникового периода. Оледенения характерны для четвертичного периода, начавшегося более миллиона лет назад. В нашей стране сохранились мерзлотные толщи, не оттаявшие даже в климатических условиях последующего этапа четвертичного периода – продолжающегося и сегодня голоцена, начавшегося 20 тыс. лет назад.
Похолодания, особенно в малоснежные зимы, вызывают продвижение на юг островов и массивов многолетней мерзлоты, и ее зона распространилась почти на все леса Восточной Сибири. На западе в европейской тайге встречаются только острова мерзлого торфа небольшой мощности (глубиной 5-10м). Возникновение на равнине водоупорного слоя многолетней мерзлоты или замерзшего торфа вызывает или усиливает заболачивание. Многолетняя мерзлота приводит к исчезновению древесных, кустарниковых и травянистых растений.
При понижении температуры нижних горизонтов почвы корни сосны начинают расти в направлении к верхним теплым горизонтам. Уменьшается прирост надземной части дерева, хвоя становится короче. Сокращается вегетационный период. Корни сосны не доходят в слои почвы с максимальной температурой ниже 3-40С, а корни ели и лиственницы проникают в слои с температурой 1-20С, то есть находятся под многолетней мерзлотой, где температура почвы колеблется от 0,2 до 3,0 0С.
Недостаток тепла в почве обедняет состав и ослабляет жизнедеятельность микроорганизмов, замедляя физико-химическое выветривание горной породы, поэтому почвы формируются поверхностные.
На юге лесной зоны в Восточной Сибири многолетняя мерзлота способствует выживанию леса в конкурентной борьбе с растительностью открытых мест. Сплошная рубка может привести к исчезновению влажного водоупорного мерзлотного горизонта, грунт иссушается, и лес сменяется почти пустынной растительностью.
Потери плодородия лесных почв происходят вследствие заболачивания, токсикоза почвы, действия кислых осадков, эрозии и пожаров.
Заболачивание почвы в таежной зоне европейской части России происходит из-за прохладного и влажного климата. При применении древостоя заболачивание может принять интенсивный характер, и заболоченный лес в слабосточных понижениях через несколько поколений превращается в болото. Примером катастрофического заболачивания является Западная Сибирь – дно широкого водоема, соединившего в далеком прошлом полярный и тропические океаны. Европейской тайге с развитым рельефом подобное не угрожает.
Процессы заболачивания в европейских лесах страны объясняются так: избыток влаги вытесняет кислород из корнеобитаемого горизонта почвы. В нем развиваются восстановительные процессы, образуются закисные соединения, происходит оглеение грунта, который становится менее водопрочным. Это подавляет жизнедеятельность почвенной микрофлоры и связанные с ней биохимические процессы гумификации и минерализации органического вещества. Неразложившиеся растительные остатки накапливаются. В результате уменьшается количество подвижных питательных веществ, ослабляется дыхательная и сосущая функции корней, нарушается функционирование фотосинтетического аппарата. В связи с этим уменьшается возврат в почву азота и зольных элементов с отмирающими частями растений. Заболачивание лесов происходит и при подтоплении от водохранилищ.
Лесные пожары приводят к заболачиванию почвы вследствие водоупорного горизонта из продуктов горения.
После сплошных рубок заболачивание почвы усиливается, и потери плодородия возрастают. Из-за переувлажнения почвы при недостатке
кислорода в почве накапливаются соединения в недоокисленной форме токсичные для растений.
Токсикоз почвы может возникнуть и при усилении подзолообразования. Накопление органических кислот способствует появлению в почвенном растворе ионов алюминия в таких концентрациях, которые вредны для растений.
Среди продуктов жизнедеятельности сапрофагов могут быть ингибиторы, которые подавляют рост и развитие растений.
Одной из причин почвенного утомления при длительном произрастании одного вида является накопление аллелопатически активных веществ (колинов), выделяемых корнями.
В составе колинов в лесной почве выделены абсцизовая кислота, флоризин, эскулин, эскулатин, фраксин, синильная кислота, флороглюцин и др. Эти вещества могут накапливаться в почве, вызывая ее переутомление.
Плодородие почвы ухудшается под влиянием кислых осадков. Многие промышленные газы превращаются при взаимодействии с парами в кислоты. Кислотный дождь имеет рН = 2,5-5,5 и содержит серную, азотную, а иногда и соляную кислоту. Лесные почвы от таких осадков становятся более кислыми, усиливается процесс выветривания грунтов. Первые симптомы поражения проявляются в накоплении в растениях напочвенного покрова Al, Fe, Na, N, Cl, Pb и др. Частично усыхает хвоя, и кроны деревьев изреживаются. Замедляется разложение подстилки, что повышает кислотность почвы.
Частичное отмирание корней активизирует деятельность стволовых вредителей, ослабляет устойчивость деревьев к засухе, морозам и лес постепенно погибает.
Отрицательно сказываются на состоянии почвы сплошные рубки. На склонах крутизной более 100 ливень смывает слой мелкозема, который образовывался 200-1000 лет.
Особенно обедняется почва после сплошных рубок в горах. Там наряду с плоскостной эрозией отмечается и внутрипочвенная.
Почвы обедняются и при систематическом древопользовании по низким оборотам рубки. При сплошной рубке с трелевкой деревьев с 1 га вывозится азота 250-400 кг, калия – 250-300 кг, фосфора 120-150 кг. Потери азота восстанавливаются через 50 лет, а потери кальция и магния не компенсируются. Поэтому американское общество по изучению лесной продукции рекомендует производить сплошные рубки после листопада, а раздробленные порубочные остатки распределить равномерно по вырубке.
При корчевании пней происходят потери плодородия почвы из-за ухудшения ее физических свойств: снижения скважности и водопроницаемости.
Лесные пожары, уничтожающие лес полностью ухудшают лесорастительные условия. Во время пожаров азот переходит в основном в газообразное состояние и до 90% его улетучивается в атмосферу. Только некоторая часть азота переходит в золу. Потоками горячего воздуха уносится до 20-40 % зольных элементов.
1. Расскажите о значении почвы в жизни леса.
2. назовите последовательно звенья биологического круговорота веществ в лесу.
3. Как влияет рельеф на компоненты леса?
4. Под влиянием каких факторов снижается плодородие почвы?
5. В чем заключаются почвоулучшающие свойства отдельных древесных пород и кустарников?
Основная литература:
Мелехов И.С. Лесоведение. М.: МГУЛ.- 2004, 398 с.
Дополнительная литература:
Погребняк П.С. Общее лесоводство. Колос. М.: 1968, 440 с.
1 Дифференциация деревьев в лесу и естественное изреживание
2 Естественный и искусственный отбор
3 Рост одновозрастных чистых древостоев различного происхождения
4 Стадии развития одновозрастных насаждений семенного происхождения
5 Возникновение и развитие разновозрастных насаждений
6 Биотические факторы в жизни леса
К возрасту главной рубки густота древостоя исчисляется сотнями деревьев на 1 га.
Саморегуляция, создающая устойчивость популяций к неблагоприятным условиям, проявляется полнее в густых древостоях. С возрастом густота уменьшается. Уменьшение количества деревьев в древостое с увеличением его возраста в результате естественного отмирания их называется естественным изреживанием древостоя. Г.Ф. Морозов назвал этот процесс законом естественного изреживания.
Естественному изреживанию древостоя обычно предшествует постепенное отставание в росте многих деревьев. Поэтому даже в одновозрастном чистом древостое в одном типе леса деревья никогда не имеют одинаковых размеров. Такое расчленение деревьев в древостое по росту и развитию его при формировании с возрастом называется дифференциацией деревьев.
В соответствии с классификацией Крафта все деревья разбиваются по степени развитости на две группы. Первая включает нормально развитые деревья (господствующие), вторая – деревья плохо развитые, отставшие в росте, заглушенные (подчиненные – по Крафту). Основными признаками для отнесения дерева к той или другой группе служат характер кроны, сравнительная высота (рост) дерева, положение среди соседних деревьев.
Одним из важных признаков является крона.
Крафт, пользуясь указанными признаками, разбивает все деревья на 5 классов (рисунок 3).
|
Рисунок 3 – Классификация лесных деревьев (по Крафту) |
I класс – прегосподствующие деревья, имеющие мощную развитую крону и крупные по высоте и диаметру стволы, выделяющиеся из общего полога. Они составляют до 8% по густоте и до 20% по запасу.
II класс – господствующие деревья, составляющие верхнюю часть древесного полога. Их насчитывается от 15 до 35%, а по запасу они составляют 40-80%. Максимальное число таких деревьев бывает в спелых древостоях, а в неспелых – после рубок ухода.
III класс – согосподствующие деревья, которые входят в общий полог с деревьями первых классов, но угнетены ими, о чем можно судить по узким кронам. Высота их меньше, количество различное: от 10% в спелых ухоженных древостоях до половины общей численности в молодняках.
IY класс – угнетенные деревья, составляющие по густоте от 5 до 30%. Делятся на два подкласса IYа – деревья с симметричной кроной, растущие в просветах, и IYб – деревья с флагообразными однобокими кронами, частично находящимися под кронами других деревьев.
Y класс – затененные деревья (от 1 до 30% по густоте). Они тоже делятся на два подкласса: Yа – деревья с жизнеспособными кронами и Yб – деревья с отмирающими или отмершими кронами.
Причинами дифференциации деревьев являются:
1. Наследственные свойства.
2. Неоднородность плодородия почвы.
3. Процесс дифференциации деревьев протекает тем интенсивнее, чем гуще естественный древостой и чем быстрее растут деревья.
4. Ослабление роста деревьев может происходить под влиянием вредителей, болезней, действия ветра, снега и другим образом.
Наиболее резко выражена дифференциация в биогруппах. Биогруппой называют близко растущие деревья, образующие непрерывный древесный полог, то есть расстояние между ними меньше среднего расстояния в древостое, его рассчитывают по формуле L = 10000/0,866·N, где N – густота древостоя.
Образование биогрупп происходит при равномерной посадке. Турский заметил, что некоторые группы деревьев остаются в том же соседстве с ростом, другие, несмотря на просторное размещение плохо прирастают. Поэтому многие считают, что биогруппы – это мощные эдифицирующие центры в лесу и управлять лесом нужно через них.
Биогруппы необходимо сохранять при рубках ухода, так как самые крупные деревья находятся в густых биогруппах. В биогруппах раньше происходит дифференциация деревьев.
В результате дифференциации возрастает влияние на древостой со стороны крупных деревьев. У отставших деревьев особенно плохо развивается корневая система, крона имеет небольшой объем. Конкурентные взаимоотношения и «ограничения» неблагоприятны для большинства растений, которые угнетены и погибают. В результате отпада слабых деревьев увеличивается площадь питания соседних, они начинают быстрее расти и устойчивость в целом повышается.
Интенсивное естественное изреживание наблюдается в сомкнутых молодняках. Потом отпад ослабляется, особенно после 70 лет. К возрасту главной рубки остается 1-5% стволов от общей густоты молодого древостоя.
Влесных культурах могут быть отклонения от общих закономерностей естественного изреживания.
Влесных культурах могут быть отклонения от общих закономерностей естественного изреживания.
Годичный отпад, не превышающий по количеству деревьев 3-10 % от общей густоты и 1-5% от объема стволовой древесины является естественным.
В естественном изреживании проявляется сущность естественного отбора и эволюции древесных пород в целом.
Положительные свойства естественного изреживания заключаются в том, что оставшиеся деревья первого яруса повышают свою жизнеспособность и имеют возможность оставить после себя семенное потомство. При этом благодаря перекрестному опылению усиливается рекомбинация генов, возрастает генетическая изменчивость, получают распространение новые, более устойчивые к изменившейся окружающей среде популяции.
Воптимальных условиях формируется более гомозиготная популяция.
В экстремальных условиях полнота древостоя небольшая, и процесс естественного изреживания замедляется, поэтому более высока вероятность выживания и плодоношения особей с иными генетическими свойствами. Частота хромосомных мутаций выше в экстремальных условиях среды. Чтобы сохранить богатство генотипов, в этих условиях нельзя проводить сплошные рубки.
Совершенствование вида не происходит при порослевом лесовозобновлении потому, что, во-первых, от пней крупных деревьев поросль не появляется или образуется в небольшом количестве, а основное число новых экземпляров – это потомство отставших в росте деревьев. Порослевые древостои оказываются менее устойчивыми при изменении условий среды.
Особенно высокая жизнеспособность и продуктивность наблюдается у экземпляров, возникших в результате скрещивания отдаленных форм. Усиленное развитие такого гибрида получило название гетерозиса. Но так как даже в первом поколении гибрида происходит сильное расщепление то ценные свойства гетерозисных особей можно сохранить только при их вегетативном размножении.
Таким образом, в лесоводстве перспективно и вегетативное размножение лесообразователей.
С наступлением вегетационного периода деревья начинают расти одновременно в высоту и по диаметру ствола, а также в подземной части.
Рост ствола по диаметру прекращается у лиственных пород в августесентябре, а у ели и сосны позже и может наблюдаться в первой декаде октября.
Рост корней продолжается дольше, чем побегов. Корни растут быстрее ночью. Максимальный рост корней в длину происходит при температуре почвы 14-190С.
Впервую очередь в течение вегетационного периода нарастает ассимиляционный аппарат деревьев, затем наблюдается максимум прироста
ввысоту, потом по диаметру ствола и в более поздние сроки идет преимущественно нарастание корней в толщину.
Рост корней и ствола зависит от климатических условий текущего и прошлого года и от уровня семеношения.
О росте древостоя в высоту и приросте по диаметру судят по изменению параметров среднего дерева. В течение жизни древостоя обычно происходит переход деревьев из высших классов Крафта в низшие.
Большинство крупных деревьев могут сохраняться в верхней части полога до возраста спелости, поэтому динамика верхнего яруса древостоя почти такая же, как у реального дерева.
В молодых древостоях более быстрыми темпами идет прирост в высоту. В 10-40-летнем древостое наступает его максимум и к концу жизни древостой прекращает рост в высоту. Заметное падение прироста происходит обычно при высоте стволов 10-12 м, когда начинается плодоношение и усиливается отрицательное воздействие ветра – раскачивание, сопряженное с началом ветровала. Прекращение роста в старом возрасте можно объяснить осмотическим ограничением подъема воды у конкретного вида в определенных условиях.
Максимальный прирост по диаметру отмечается одновременно или на 5-15 лет позже максимального прироста по высоте. Через 10-30 лет после максимума прироста по диаметру наблюдается максимум накопленного запаса древесины. При этом сомкнутость древесного полога максимальна (0,95) в 10-40 летнем возрасте. Максимальная производительность наблюдается в древостоях с перекрытием крон 25-40%. Это указывает на целесообразность разреживаний перегущенных чистых древостоев рубками ухода.
Чем выше класс бонитета, тем раньше наступает кульминация прироста.
Вперегущенных древостоях прирост по высоте и диаметру снижается из-за недостатка влаги, света, минеральных веществ и азотистых соединений.
Некоторыми особенностями роста характеризуются древостои вегетативного происхождения. Рассмотрим на примере порослевых дубрав. Посте сплошной рубки дубравы образующаяся пневая поросль растет быстрее дубков семенного происхождения благодаря развитой материнской корневой системы. Из-за недостатка поверхности листвы молодняков, корневая система постепенно отмирает. Через 2-3 года у поросли ниже корневой шейки образуются придаточные корни, которые не достигают размеров материнских корней. Поросль имеет максимальный прирост в высоту в первые 5 лет. Дубки семенного происхождения догоняют по высоте порослевые особи во 2-4 десятилетии, в период интенсивного роста, тогда как порослевые деревья все больше снижают темпы роста. С каждой генерацией порослевого поколения уменьшается масса активных корней, рост в высоту ослабляется, деревья деградируют.
В порослевых дубравах кроны разрастаются в большей мере, так как расстояние между деревьями смежных гнезд больше, чем в семенных молодняках. Усиливается сучковатость, кривизна стволов. Они поражаются гнилью от распадающейся материнской корневой системы.
Древостои растут всю жизнь, а развитие их происходит по этапам. 1.Стадия индивидуального роста характеризуется неблагоприятными
условиями существования древесных растений. Самосев и культуры страдают от ветра, прямой солнечной радиации, иссушения почвы. На этой стадии их особи гибнут от опала корневой шейки, заморозков, выжимания кристаллами льда, вымокания и других причин. В этот период ведущими являются межвидовые отношения. Эта стадия продолжается от 3 до 15 лет.
2. Стадия чащи. На этой стадии древостой имеет не только максимальную густоту, но и максимальную сомкнутость крон, отмечается наивысшая динамика состава древостоя в связи с обострением межвидовых отношений среди древесных пород. Чтобы она не изменилась в нежелательную для хозяйства сторону, требуется проведение рубок ухода.
Стадия чащи продолжается до возраста древостоя 10-25 лет. Затем возросшая мощность древесного полога, задерживающего свет и корневая конкуренции приводят к изреживанию травяного яруса. Древостой достигает высоты 6 м и выше, быстро растет в высоту. Поэтому следующая стадия называется жердняком или стадия усиленного роста.
3.Стадия усиленного роста (жердняка) является вторым критическим периодом в жизни популяции, потому что в древостое наступает максимум прироста по высоте, наблюдается повышение прироста ветвей и листьев.
С этой стадии развития насаждения начинается полное влияние леса на почву, температуру воздуха и его движение под пологом, восстанавливаются защитные функции леса.
4.Стадия возмужалости характеризуется началом систематического плодоношения деревьев, которое наблюдается у различных пород с 20-60 лет. На этой стадии отмечается максимум транспирации, прекращается прирост в глубину вертикальных ответвлений корней господствующих деревьев.
Со временем начинают отпадать не только отставшие в росте деревья, но и средние и средние и крупные у которых быстро прошел период интенсивного роста, и раньше наступило старение. В образовавшихся «окнах» появляется подрост. Происходит переход к следующей стадии.
5.Стадия предварительного возобновления наиболее продолжительна.
Во время этой стадии снижаются темпы потребления веществ, а возраст растет. Из подроста хозяйственно ценных пород образуется новое поколение леса. В древостоях из светолюбивых древесных пород, находящихся в стадии предварительного возобновления, может образоваться подрост теневыносливых пород.
На смену старому поколению формируется новое поколение леса.
6. Стадия смены поколений нежелательна в хозяйственном отношении. М.К. Турский назвал момент уменьшения запаса древостоя возрастом естественной спелости. До его наступления одновозрастные насаждения должны поступить в главную рубку. Стадия смены поколений продолжается 60-80 и более лет, в течение которых происходит отпад старых деревьев. На протяжении этой стадии общий запас древостоя уменьшается. Постепенно подрастает второй ярус, и наступает момент нового роста запаса. Так формируется типичный разновозрастный древостой.
По различиям в происхождении древостоев и неодинаковым экологическим свойствам пород выделяют несколько типов возрастных структур лесов. Наиболее характерны из них: 1.) абсолютно разновозрастные древостои, образующиеся в результате выборочных рубок или при длительном развитии лесов из теневыносливых пород;
2.) относительно разновозрастные древостои, основное число деревьев которых появилось в течение 60 лет, и ряд распределения деревьев по толщине похож на ряд одновозрастных насаждений, но очень растянут;
3.) промежуточные типы возрастных структур, в которых легко можно выделить поколения леса, а ряды распределения деревьев по толщине имеют несколько вершин.
Новое поколение леса в разновозрастном древостое также проходит ряд возрастных стадий, отражающих онтогенез этого поколения.
1 стадия возобновительная от 5 до 40 лет. К концу ее накапливается 2- 6 тыс. экземпляров подроста на 1 га их кроны смыкаются. При достижении сомкнутости молодого полога 0,4 особи начинают влиять друг на друга, и поколение вступает во 2 стадию – угнетенного яруса, где особи подвергаются конкуренции, что приводит их к опаду.
По мере уменьшения числа деревьев старого поколения увеличивается прирост в высоту нового поколения и наступает стадия большого роста. Она продолжается до 60 лет.
Окончательный распад старого поколения или рубка его деревьев вызывают не только интенсивный рост нижнего яруса в высоту. Отдельные деревья получают полное освещение и начинают плодоносить, что указывает на начало следующей стадии возмужалости. Характерным для нее является выраженная дифференциация деревьев и отпад тонкомерных экземпляров.Последующая стадия зрелости характеризуется снижением устойчивости деревьев, ослаблением прироста в высоту и максимальным приростом в толщину.
Последняя стадия – распад поколения – начинается с момента уменьшения ее запаса (у сосны со 180 до 320 лет).
Одновозрастные древостои характеризуются обычно горизонтальной, разновозрастные – вертикальной сомкнутостью полога. Между этими двумя видами возрастной структуры имеется ряд промежуточных, которые лишь условно могут относится или к одновозрастной или к разновозрастной структуре (рисунок 4).
|
Рисунок 4 – Схема древостоев а – соснового одновозрастного; б – елового разновозрастного; |
Таким образом, разновозрастные леса являются одновременно и старыми и молодыми. Разновозрастные древостои преобладают в нашей стране.
Типы влияния растений друг на друга
Н.В. Сукачев объединил примеры взаимоотношений между растениями в 3 группы случаев:
1.Оба или несколько видов лучше развиваются в смеси, чем в чистых посевах – встречается редко.
2.Из двух видов один лучше чувствует себя в смешанном, другой – в числом посеве – встречается часто.
3.В смеси оба вида чувствуют себя плохо, что связано с взаимно
неблагоприятными выделениями коневых систем. (не часто) Взаимоотношения растения имеют многообразный характер. Выделяют следующие типы влияния растений друг на друга:
1.механическое, выражается в виде охлестывания вершин ветвями другого дерева;
2.биофизическое влияние путем изменения факторов среды (света, тепла, влаги, пищи);
3.биохимическое или аллелопатическое – воздействие биологически активных веществ (фитонцидов), выделяемых различными органами;
4.биотрофное взаимодействие, происходящее в зоне ризосферы в процессе потребления и выделения элементов питания;
5.физиологическое взаимодействие – срастание корней, паразитизм, симбиоз;
6.генеалогическое влияние, наблюдается при опылении цветков и зачатков растений.
При описании связей древесных пород с абиотическими факторами среды мы рассматривали основной компонент фитоценоза – древостой и его связи с почвенным микробоценозом и почвенной фауной. Экология леса была бы неполной, если не рассмотреть остальные части биогеоценоза: микробоценоз верхних биогоризонтов, растительность нижних ярусов и зооценоз.
Постоянно функционирующим живым элементом почвы является микробоценоз. Хотя составляющие его организмы (бактерии, грибы, водоросли) встречаются и в верхних биогоризонтах, но основная их масса (0,5-1.0 т/га) находится в почве. Они не только разрушают органические остатки, минералы, гумус, но и создают новые вещества. Энергию роста и размножения микроорганизмы получают при окислении (аэробы) и брожении (анаэробы) различных веществ. Микроорганизмы лучше развиваются при нейтральной реакции почвенного раствора, но некоторые приспособились переносить кислые почвы (плесени) или щелочные (нитрофикаторы).
Для большинства микроорганизмов оптимальной температурой является 25-300С, а наилучшая влажность почвы та же что и для деревьев. Микроорганизмы устойчивы, способны жить в зимних условиях.
Взаимоотношения между компонентами фитоценоза
Древесные породы влияют не только друг на друга, но и на нижние ярусы растительности. Наибольшему влиянию подвержен живой напочвенный покров. Опад деревьев может содержать вещества, тормозящие прорастание семян травянистых растений.
Травянистый ярус является защитой и пищей животных, разрыхляет почву, обогащает ее органическим веществом, улучшает структуру и таким образом влияет на течение дернового процесса.
Подлесок также играет положительную роль. Благодаря его опаду происходит обогащение верхних горизонтов почвы азотом и зольными элементами, снижается кислотность почвы. Подлесок ослабляет эрозию почвы. Наличие подлеска повышает численность орнитофаюны и устойчивость против вредных насекомых. Особенно велика положительная роль подлеска на опушках леса.
Кустарники оказывают аллелопатическое влияние на древесные породы. Аллелопатия – прямое химическое воздействие растений друг на друга. Они могут носить положительный характер, когда растение выделяет через листья или корни вещества, стимулирующие рост и развитие соседа. Чаще устанавливаются отрицательные взаимоотношения, при которых выделения растений носят так называемый ингибиторный характер – задерживают рост и развитие соседей вплоть до их умерщвления.
Ингибиторами березы (при совместном произрастании) является аморфа кустарниковая, жимолость татарская, бузина красная, смородина золотая, терн. Активаторами – лох узколистный, акация желтая, ирга.
Отрицательная роль подлеска может проявиться при возобновлении древесных пород под пологом леса.
Подрост дуба часто угнетается лещиной.
Среди представителей внеярусной растительности особое место занимает омела – полупаразит, вызывающий при сильном развитии засыхание ветвей и вершин деревьев.
На иве, тополе, ольхе паразитируют 3 вида повилики, которые могут причинять вред только молодым деревьям.
Незначительную роль в жизни лесного биогеоценоза играют эпифиты (от греческого epi – на и phytonрастение), представленные в лесах умеренной зоны мхами, лишайниками, водорослями, поселяющимися на стволах и ветвях деревьев. Они не проникают в ткани деревьев, являются автотрофными организмами, самостоятельно синтезируют органические вещества и растут в основном весной и осенью. Воздушно-сухая масса их может достигать 1-2 тонн на 1 га.
Некоторые представители фитоценоза имеют узкую экологическую нишу, тесно связаны с определенными свойствами почв и получили название растений-индикаторов. Так, лишайники, вереск – встречаются на бедных сухих почвах, клюква, морошка – на торфяниках, кукушкин лен обыкновенный – на сырых бедных почвах, вейник ланцетный – на средних по плодородию, а таволга и звездчатка лесная – на богатых почвах. Во влажных лесорастительных условиях растут – черника, щитовник игольчатый, сныть, крапива, на свежих бедных почвах – зеленые мхи, брусника; на относительно богатых – звездчатка, зеленчук желтый, а на богатых – медуница, осока волосистая.
Роль фауны в лесном биогеоценозе
В отличие от других компонентов леса многие виды животных не присущи только конкретному биогеоценозу, их местообитание (стация) распространяется на целые ландшафты. Эти виды называются евритопными,
вотличие от стенотопных животных, жизнь которых приурочена к одному или ограниченному числу лесных биогеоценозов.
Всостав зооценоза входят млекопитающие, птицы и др. позвоночные и беспозвоночные животные.
Основное влияние фауны оказывает на древостой, в частности на семенное возобновление. Некоторые насекомые воздействуют на цветочные почки. Хвое и листогрызущие насекомые ослабляют возможность образования генеративных органов. Цветочные почки могут быть съедены белкой, рябчиком. Молодые шишки сосны склевывают тетерева и глухари. Созревшие семена съедают кабаны, белки, полевки и др.
Положительная роль насекомых проявляется при опылении цветков. Некоторые древесные породы (липа, клен, черемуха) являются энтомофилами. Многие животные и птицы являются распространителями семян (сойка, кедровка).
Появившиеся всходы скусывают муравьи, полевки, мыши, а стволики саженцев подроста обгладывают зайцы, корни деревьев объедают личинки майского хруща и повреждают медведки и др.
Однако у вредителей есть враги. Использование естественного антагонизма различных видов составляет суть биологических мер борьбы с вредителями. Один из методов – не создавать благоприятных условий для размножения вредителей. От вредителей чаще страдают древостои, расстроенные бессистемными рубками, ветром, пожарами, болезнями, загрязнением атмосферы и высокой численностью отдыхающих.
Некоторые дендрофильные насекомые в нарушенных биогеоценозах становятся стволовыми вредителями.
В нормально развивающихся коренных естественных древостоях каждое дендрофильное насекомое имеет свою экологическую нишу, поселяясь на ослабленных деревьях определенной категории.
Птицы и млекопитающие оказывают непосредственное влияние на почву тем, что перемешивают лесную подстилку с ее минеральной частью. Разрыхляют почву и удобряют ее пищевыми остатками, экскрементами.
Почвенные животные, прокладывая ходы, портят корни травянистых растений, но, выбрасывая землю на поверхность, способствуют развитию на ней всходов древесных пород.
Таким образом, каждый вид лесной фауны имеет свое значение в жизни леса. Если видов много, плотность населения каждого из них сужается, и снижается опасность вспышки численности вредителя.
Изменение численности одного вида приводит к изменению количества особей других видов, связанных с ним пищевой цепью.
Значение пищевых цепей и оптимального соотношения числа особей каждого вида в них (включая и растительность) является необходимым условием поддержания в лесу биологического равновесия.
Разнообразие и численность животных зависят от кормовых ресурсов. К.С. Элтон построил трофическую пирамиду, состоящую из 4-5 трофических уровней. Нижний уровень – первичная продукция, то есть масса автотрофных растений – продуцентов, которая является кормом для растительноядных животных – фитофагов, или консументов, заполняющих второй уровень. Он имеет меньшую массу (почти в 10 раз) и, являясь пищей для плотоядных организмов 1 порядка (3 уровня), а последние служат пищевым ресурсом для консументов высшего порядка (четвертого уровня) и т. д.
Последний трофический уровень имеет наименьшую биомассу и венчает пирамиду Элтона.
Хищник каждого последующего трофического уровня обычно больше по размеру, чем предыдущего, поэтому численность его должна быть меньше.
Биомасса подземных представителей животного мира выше биомассы надземных животных. Тля – вредное насекомое, однако выделяемая ею медвяная роса служит источником питания муравьям. Муравьи улучшают водно-воздушный режим почв и обогащают ее гумусом. Они уничтожают хвое и листогрызущих вредителей. В свою очередь они являются пищей позвоночных животных – птиц.
Хищники могут уничтожить в пищевой цепи не более половины массы животных низшего уровня.
Особое звено пищевой цепи составляют беспозвоночные, съедающие животный опад. На трупах млекопитающих существуют жуки – мертвоеды, карапузики, личинки мух, гусеницы бабочек – керафагов, поедающие шерсть и рога.
Своеобразную пищевую цепь составляют паразитические микроорганизмы, предотвращающие эпизоотии своих хозяев фитофагов.
Типы взаимоотношений в лесном биогеоценозе
В лесном биогеоценозе проявляются самые разнообразные взаимоотношения между особями и популяциями животных и растений. Различают следующие типы общебиологических взаимоотношений: хищничество, паразитизм, конкуренция, ограничение, комменсализм и мутации (симбиоз).
Хищничество – очень распространено в лесном биогеоценозе и заключается в ловле и поедании представителя одного вида особей другими. По отношению к травянистым растениям растительноядные животные должны считаться хищниками. Среди многочисленных примеров хищничества представляет интерес поедание мелких насекомых растениемросянкой круглолистной.
Паразитизм – это способ существования одного вида (паразита), за счет другого (хозяина). Он может не вызывать гибель используемого организма и наблюдается между растениями и животными (например, насекомые – вредители), животными и микроорганизмами.
Известен паразитизм и между растениями. К паразитизму относят разрушение ствола гнилями грибов.
Ограничение – это пропорциональное потребление соседними особями веществ и энергии при их недостатке, которое в перегущенных культурах снижает прирост стволовой древесины и может привести к гибели лесов.
Конкуренция способствует выделению лидеров и завершается естественным отпадом более слабых особей.
Комменсализм – симбиотические взаимоотношения между двумя видами, когда один вид питается за счет другого, не нанося ему ущерба. Примером служит жизнь эпифитов на стволах деревьев.
Мутуализм или симбиоз. Это когда совместное существование выгодно только для одного вида и безразлична для другого.
Регулирование плотности населения дикой фауны
Плотность населения крупных животных принято показывать в расчете на 1000 га, птиц – на 10 га. Плотность населения лесных видов обычно относят на единицу покрытой лесом площади.
Численность фауны обусловлена состоянием кормовой базы, развитием болезней, миграциями и интенсивностью промысла. В условиях ограниченной кормообеспеченности необходимо строгое регулирование плотности населения животных, в противном случае они могут причинять большой ущерб лесному хозяйству.
Вопросы регулирования численности фауны представляют интерес как для лесного, так и для охотничьего хозяйства.
По данным института «Союзгипролесхоз» (в 70-е годы) для охотничьих угодий третьего бонитета (т. е. среднепроизводительные) численность фауны должна быть следующей (шт. на 1 тыс. га) : лось – 5, олень – 10, косуля – 40,кабан – 8, заяц-беляк- 55, заяц русак – 30, куропатка серая – 150, тетерев – 100, глухарь – 40.
Экологическая роль пастьбы скота в лесу Лесные площади нередко используются для пастьбы скота. Пастьба
скота, особенно неумеренная, приносит вред лесу – разрушается подстилка, легкие почвы разбиваются, иссушаются, песок развеивается ветром, и тяжелые почвы уплотняются. На таких почвах ослабляется дыхание корней из-за недостатка кислорода, они загнивают, тонкие корни выжимаются кристаллами льда. Обнаженные корни повреждаются копытами, в местах повреждений появляются гнили.
Снижается и качество древесины, так как скусывание вершин приводит к кривоствольности, а обгладывание коры вызывает сухобокость и развитие стволовой гнили. Особенно опасны в этом отношении козы, пастьба которых в лесу недопустима.
Пастьба скота изменяет состав живого напочвенного покрова. Теневыносливые лесные виды сменяются луговыми, происходит задернение почвы, что затрудняет развитие всходов. В целом процесс естественного лесовозобновления, особенно главных пород, ухудшается как под пологом леса, так и на вырубках.
Пастьбу скота следует допускать только на территории, которая со временем будет преобразована в высокопродуктивные сельскохозяйственные угодья.
1. Укажите основные причины естественного изреживания и дифференциации деревьев в лесу.
2. Расскажите о классификациях деревьев в лесу.
3. Какие вы знаете стадии развития одновозрастных насаждений?
4. Какими путями регулируется плотность населения дикой фауны?
5. Какие вы знаете типы взаимоотношений в лесном биогеоценозе?
6. Каковы взаимоотношения между компонентами фитоценозов?
Мелехов И.С. Лесоведение. М.: МГУЛ.- 2004, 398 с.
Дополнительная литература:
Погребняк П.С. Общее лесоводство. Колос. М.: 1968, 440 с.
1 Истоки лесной типологии 2 Учение о типах леса Г.Ф. Морозова
3 Классификация условий произрастания П.С. Погребняка
4 Классификация типов леса В.Н. Сукачева
5 Типология степных лесов А.Л. Бельгарда
6 Особенности лесной типологии за рубежом
Идея лесной типологии зародилась в народе. А.Е. Теплоухов писал, что крестьяне называли мелкослойную, смолистую, крепкую осину брусняжной, так как почва, поросшая брусникой, указывает на древесину хороших качеств. Такие народные названия, как рамень, согра, бор, лог, раза и др. отражают единство древостоя с условиями его произрастания и выявляют типологические различия лесов.
В18 веке лесовод А.А. Нартов впервые отметил связь роста древостоя
с условиями произрастания, которые он подразделил на сухие, сырые земли и внебровочные участки с клюквой и мхом.
Вконце 19 века из-за возросшего объема сплошных рубок обострилась проблема лесовозобновления. В.Я. Добровлянский высказал мысль о необходимости учения лесовозобновления по предварительно выделенным «типам насаждений», разнообразие которых определяется, по его мнению, внешними условиями (климатом и почвенно-грунтовыми условиями), внутренними условиями, создающимися самим насаждением (составом, густотой древостоя и т.д.) и влиянием человека.
А. Ф. Рудзкий предложил насаждения разделять на небольшое число «условных однообразий» или «первообразов», принимая во внимание высоту над уровнем моря, экспедицию склона, почву, хозяйственную значимость древесной породы, густоту и возраст древостоя. Он дал пример 7 таких «первообразов» европейской тайги в т. числе: 1).сосна кондовая или боровая на свежей и глубокой почве; 2) . Сосна мяндовая на сырой почве; 3). Сосна болотная на мшарине; 4).Хвойный лиственный лес: пестрая смесь с неопределенным господством сосны, ели, березы, осины и др.
И. И. Гуторович при лесоустройстве Вологодской губернии описал 9 «типов насаждений», приняв во внимание 4 фактора: топографическое положение участка, почву, состав насаждения и напочвенный покров. Некоторые из них близки к современным типам леса: «рада» – сосняк сфагновый, «суболоть»- сосняк багульниковый, «согра» - сосняк (ельник) осоковосфагновый, «биль»- близок к сосняку (ельнику) черничному, «бор»
– близок к сосняку брусничному, «ровнядь» - ельник долгомошный, «холм»- ельник кисличный, «лог»- ельник - лог.
Вконце 19 века проявляли интерес к типологии лесов и ботаники. С. И. Коржинский считал, что в результате длительной борьбы за существование в каждом участке с присущими ему климатическими и топографическими условиями сформировалась определенная совокупность видов растений – ассоциация. Ассоциации объединялись в более устойчивые «общежития»- растительной формации. Поэтому Коржинский рассматривал типы насаждений, как типы растительных формаций. Им была вскрыта роль растений - эдификаторов, которые определяли внутреннюю среду сообщества и его развитие.
Необходимость деления лесов на однородные единицы (типы) насаждений впервые научно обосновал Г. Ф. Морозов. При установлении типов на раннем этапе он принимал во внимание условия произрастания, а в дальнейшем целую совокупность признаков:
1). внутренние, экологические свойства древесных пород; 2). географическую среду - климат, грунт, рельеф, почву; 3). биосоциальные отношения:
а) между растениями, образующими лесное сообщество; б). их же со средой; в). их же с фауной;
4).историко-географические причины;
5). вмешательство человека.
В 1904 году Г. Ф. Морозов писал: «Тип насаждения есть совокупность насаждений, объединенных в одну обширную группу общностью условий местопроизрастания, или почвенно-грунтовых условий. Он не создал законченной классификации типов насаждений, но для отдельных лесных массивов России привел примеры лесоводственнотипологического описания боров и дубрав. Идея типов леса Г. Ф. Морозова послужила основой для развития двух типологических направлений.
Опираясь на более ранние работы Г. Ф. Морозова, А. А. Крюденер разработал условия местопроизрастания с учетом рельефа, влажности и механического состава. Дальнейшее развитие это направление получило в 20х годах текущего столетия в трудах Е. В. Алексеева, П. С. Погребняка,
Д. В. Воробьева. Тип лесорастительных условий для покрытых и не покрытых лесом площадей они определяли с учетом плодородия и влажности почвы. Все разнообразие выделенных типов леса П. С. Погребняк разместил в эдафической сетке (рисунок 5).
П.С. Погребняк объединил типы леса на суходолах и в мокрых местообитаниях в один трофогенный ряд. Совместно с Д. В. Воробьевым они выделили 5 трофотопов ( А - боры, В - субори, С- сугрудки , Д - груды, Е- дубравы), затем осталось 4. В настоящее время термин «дубрава» используют синонимы: рамени (в ареале ели); бучины (в ареале бука), а для термина «сложные субори» - сугрудки, сурамени, судубравы, субучины.
|
Рисунок 5 – Эдафическая сетка лесов по П.С. Погребняку |
Называя почвы по гранулометрическому составу корненасыщенного верхнего горизонта, П. С. Погребняк установил, что песчаные почвы могут быть самого различного плодородия от боров (А) до дубрав (Д).
К борам относятся однородные пески, без каких либо прослоек, до гл. 2,5-3 м. При наличии редко чередующихся тонких (0,5-1 см) супесчаноглинистых прослоек глубокие пески относятся к простым суборям (В). Сложные субори (С) под песками имеют на глубине не < 0,5 м богатую
почвообразующую породу. При дальнейшем уменьшении мощности песчаного наноса (0,5м и меньше), когда подстилающий суглинок интенсивно осваивается древесными корнями, почва по плодородию переходит в разряд дубравных (Д).
Но и в одном трофотопе почвы могут быть не вполне однородные по гранулометрическому составу, но однородные по богатству.
Боры (А)- крайне бедные почвенно-грунтовые условия; простые субори (В)-относительно бедные; сложные субори (С)- относительно богатые; дубравы (Д)- наиболее плодородные почвы.
Гидротопов П. С. Погребняк выделил 6.
Для большинства лесорастительных подзон самыми благоприятными являются свежие условия местопроизрастания.
Сочетания одного трофотопа с одним гигротопом дает эдатоп.
С учетом подразделений в эдафическую сетку включают все разнообразия лесов.
Применение лесотипологической классификации П. С. Погребняка в лесохозяйственной практике нашей страны способствовало улучшению ведения лесного хозяйства, изучению природы лесов, более рациональному использованию лесных земель и повышению продуктивности насаждений.
Второе лесотипологическое направление длительное время возглавлял В. Н. Сукачев. В основу классификации типов леса он положил растительные ассоциации, в дальнейшем все компоненты леса и взаимосвязь их между собой и лесорастительными условиями. Тип леса он определял по совокупности признаков, называл по растениям – индификаторам, господствующей древесной породе, преобладающему виду напочвенного покрова и устанавливал только для покрытых лесом площадей.
Под типом леса В.Н. Сукачев понимал «участки леса, однородные по составу древесных пород, по другим ярусам растительности и фауне, по комплексу лесорастительных условий (климатических, почвенных, гидрологических), по взаимоотношениям между растениями и средой, по восстановительным процессам и направлению смен в них а, следовательно, при одинаковых экономических условиях требующие однородных лесохозяйственных мероприятий».
Все разнообразие выделенных типов леса В.Н. Сукачев расположил в системе эколого-фитоценотических рядов:
Ряд А – уменьшение плодородия и влажности почвы; В – увеличение застойного увлажнения; С – увеличение богатства почвы; Д - увеличение проточного увлажнения;
Е – переходный от застойного увлажнения к проточному.
|
Рисунок 6 - Схема эколого-фитоценотических рядов сосновых лесов. |
Он предложил систему эколого-фитоценотических рядов для сосняков, ельников, и обобщенную схему эдафо-фитоценотических ареалов (таблица).
Различия и сходство типологических классификаций В. Н. Сукачева и П. С. Погребняка.
Различия между двумя основными лесотипологическими направлениями заключаются главным образом в разном объеме понятия «тип леса». Даже таксон «тип древостоя» отражает особенности географической среды и древостоя, тогда как тип леса по В. Н. Сукачеву, выделяется и по особенностям древесных компонентов леса, и по взаимоотношениям между ними. Ряды типов лесных биогеоценозов у В. Н. Сукачева не ограничены, а число типов зависит от разнообразия действия факторов лесообразования. Число типов леса у Погребняка определяется хозяйственной необходимостью.
В лесохозяйственной практике при проведении отдельных мероприятий необходимо иметь сведения о разнообразии почвенногрунтовых условий. Поэтому наряду с технологией Сукачева используется классификация Погребняка, т.е. во время наземной таксации леса при лесоустроительных работах в каждом таксационном выделе указывается наиболее распространенный тип лесного биогеоценоза и тип лесорастительных условий.
Типология В. Н. Сукачева разрабатывалась на основе изучения равнинных таежных девственных лесов нашей страны. Основным объектом являлся лес со всеми его компонентами.
|
Рисунок 7. Схема эколого-фитоценотических рядов еловых лесов. Ельники: 1 - кисличник; 2 - брусничник; -3 - черничник; |
Типология А.Л. Бельгарда была разработана в 1971 году для степных естественных и искусственных лесов.
Группу естественных лесов представляют пойменные, байрачные и аренные леса. Взяв за основу эдафическую сетку П.С. Погребняка, он заменил ординату трофности на ординату минерализованности и выделил следующие трофотопы:
АВ – физически бедные почвы, чаще всего глинистые пески; В – относительно физически бедные почвенно-грунтовые условия,
обычно связанные с легкими супесями или глинистыми песками; ВС – относительно физически бедноватые почвенно-грунтовые
условия; С – относительно богатые трофотопы, связанные с супесчаными
почвами; Д – наиболее богатые местообитания, тяготеющие к плодородным
суглинкам или супесям, подстилаемым прослойками глин или суглинков; Дсвстречается на более выщелоченных почвах; Дас – характеризует наиболее благоприятные лесорастительные
условия, способствующие формированию сложных лесных ценозов; Дп – тяготеет к почвам, отличающимся известной карбонатностью и в более влажных гигротипах – развитием нитрификационных процессов; Де – формируется в пределах продолжительнопоемных местообитаний и отличается некоторой солонцеватостью;
Е – типы леса, формирующиеся на физиологически относительно бедных почвах, которые характеризуются в поймах признаками засоления, а в условиях овражно-балочных систем - карбонатностью.
Кроме того, выделены еще трофотопы F и G, связанные с кустарниковыми ценозами.
По характеру увлажнения выделено девять гигротопов: 0 – очень сухие местообитания; 0-1 – сухие; 1 – суховатые;
1-2 – свежеватые;
2 – свежие;
2-3 – влажноватые;
3- влажные;
4 – сырые;
5 – мокрые.
Пойменные леса подразделяются на 2 группы – продолжительнопоемные и краткопоемные, применительно к которым выделены типы леса.
Типология искусственных лесов степной зоны строится А.Л. Бельгардом с учетом типов местоположения (поемные, аренные и плакорные).
В пределах типа местоположения, почвы по гранулометрическому составу делятся на: пески (П), супеси (СП) и суглинки (СГ). Засоленные почвы имеют свой индекс (З). Учитывается тип (подтип) почвы (обыкновенные черноземы, южные черноземы, темно-каштановые почвы).
Гигротопы выделяются по тем же признакам, что и для естественных лесов (0 – очень сухие; 0-1 –сухие; 1 - суховатые;1-2 – свежеватые; 2 – свежие; 3- влажные; 4 – сырые; 5 – мокрые.
Типология А.Л. Бельгарда сложна, громоздка, отличается большой дробностью, но она позволяет достаточно полно оценивать лесорастительные условия и определять как ассортимент пород для выращивания, так и тип лесного насаждения, в наибольшей степени соответствующий этим условиям.
Ученые различных стран с интересом относятся к достижениям отечественной лесной типологии. Лесоводы Польши, Болгарии, Швеции используют классификационные принципы, сходные с эдафической сеткой П.С. Погребняка.
Австрийское лесотипологическое направление Е. Айхингера, сложившееся в 50-60-е годы предлагает историческое подразделение типов леса: 1) ельники, сменившие криволесье из горной сосны, 2) ельники, сменившие леса из европейского кедра, 3) ельники, сменившие леса из лиственницы и т. д.
В зарубежной литературе местообитание рассматривается как экосистема. Каждая единица местообитания характеризуется особенностями топографии, почвы, микроклимата, состава растительности всех ярусов. Величина одного местообитания определяется темпами роста древостоя.
Впределах Баден-Вюртемберга региональная классификация включает 7 крупных лесных ландшафтов, различающихся по климату, геологии и почвам (бассейн Верхнего Рейна и др.). Они подразделяются на мелкие ландшафты, или «округа роста», не только по указанным признакам, но и по растительности с учетом особенностей древних лесов (с 1500 г. до н. э.) по пыльцевому анализу. Каждый «округ роста» подразделен на 10-15 единиц местообитания отличающихся по степени влажности и другим особенностям почвы и топографии. Единица местообитания объединяет древостои, обладающие сходным лесоводственным потенциалом и наличием или отсутствием определенных экологических групп растений.
Классификация местообитаний применяется в исследованиях по лесной селекции, почвоведению и защите леса.
1. Расскажите об истоках лесной типологии.
2. В чем состоят различия и сходства типологических классификаций В. Н. Сукачева и П. С. Погребняка?
3. Какие существуют связи типов вырубок с типами леса?
4. Расскажите о значении типологии в познании природы леса и ее использовании в практике.
5. Охарактеризуйте концепцию типологии А.Л. Бельгарда.
6. Что из зарубежного опыта вы считаете пригодным для совершенствования региональной классификации типов леса?
Основная литература:
Мелехов И.С. Лесоведение. М.: МГУЛ.- 2004, 398 с.
Бельгард А.Л. Степное лесоведение М.: «Лесная промышленность», 1971, 336 с.
Дополнительная литература:
Погребняк П.С. Общее лесоводство. Колос. М.: 1968, 440 с.
1 Методы возобновления леса
2 Виды лесовозобновления и размножения древесных пород
3 Факторы семенного возобновления
4 Классификация подроста
5 Методы изучения лесовозобновления и его оценка
6 Смена пород Причины и виды смены пород
Возобновлением леса называется образование нового поколения древесных растений.
Различают следующие методы возобновления леса: естественное, искусственное и комбинированное.
Естественное возобновление леса – это возобновление леса естественным путем. Оно рассматривается как один из методов лесовосстановления, так как это не стихийный процесс, а направляемое лесоводами явление.
Активное воздействие обеспечивается мерами содействия естественному возобновлению, а также методами проведения главных рубок и рубок ухода в молодняках. Поэтому успешность естественного возобновления зависит от квалификации лесоводов.
Естественное лесовозобновление имеет преимущество в том, что это местная популяция, ее особи произошли от материнских деревьев, сохранившихся в результате борьбы за существование на данном месте. Поэтому они оказываются более устойчивыми к местным отрицательным факторам и более приспособленными к данным лесорастительным условиям, чем лесные культуры.
Искусственное лесоразведение, по ОСТ 56-108-98, это возобновление леса созданием лесных культур.
К искусственному лесоразведению следует относить сплошные культуры, встречаемость особей которых будет>65%. В соответствии с действующей лесоустроительной инструкции древостои относят к лесным культурам, если древесные породы искусственного происхождения составляют в них не менее трех единиц от состава насаждений.
Искусственное лесовозобновление часто требует больше затрат труда и средств. Поэтому, к созданию лесных культур надо прибегать в том случае, когда естественное возобновление оказывается экономически менее эффективным.
К искусственному лесоразведению следует стремиться в том случае, когда по каким-то причинам главная порода естественным путем не возобновляется. В основном это вырубки на месте сложных и травяных типов леса.
Комбинированное - естественное и искусственное возобновления леса в различных сочетаниях на одном участке.
По времени появления особей древесных пород после проведенной на участке главной рубки различают предварительное, сопутствующее и последующее возобновления.
Предварительное возобновление лесаэто естественное возобновление под пологом древостоя. Появившиеся в результате естественного возобновления леса особи называются предварительным подростом.
Если поколение древесной породы возникло благодаря целенаправленному изреживанию древостоя для создания условий, способствующих возобновлению главной пород, то оно относится к сопутствующему возобновлению.
Сопутствующим возобновлением леса называется лесовозобновление, происходящее в насаждении в связи с рубками древостоев.
Последующим называется возобновление леса после вырубки древостоев или исчезновения их по другим причинам. В нашей стране распространено последующее возобновление естественным, искусственным и комбинированным методам.
Древесные породы размножаются семенным (бесполым) путем. Семенное размножение присуще всем нашим лесообразователям.
Естественное вегетативное возобновление может происходить порослью от пня, ствола и корней, а также отводками от корневищ. Искусственное возобновление может производиться черенками, кольями и др. путем.
Под порослевым возобновлением следует понимать только вегетативное возобновление от пня. Так размножается береза, дуб, ясень и другие хвойные за исключением тиса, практически не образуют поросль.
Размножение порослью корней называется корнеотпрысковым, а поросль от ствола - водяными побегами.
Размножение отводками имеет практическое значение у липы, рододендрона, пихты, дуба скального.
Таким образом, при анализе лесовозобновления следует выделить подрост предварительный, сопутствующий и последующий. Каждый из них может быть представлен самосевом и особям вегетативного происхождения. Поросль может относиться к сопутствующему или к последующему подросту.
В отличие от корнеотпрыскового возобновления, размножение порослью от пня зависит от возраста древостоя и диаметра пней.
Большинство побегов образуется на пнях от деревьев семенного происхождения, чем от порослевых особей.
В конце 19 века было доказано, что лесовозобновление является многофакторным процессом. Оно зависит от биологических и экологических свойств древесных пород, обуславливающих семеношение и конкурентные взаимоотношения с другими растениями, от наличия тепла, влаги, света, мощности лесной подстилки и ее биохимических свойств, от захламленности территории, от влияния других растений, животного мира, болезней, огня и так далее.
Биологические свойства обсеменителей
Мужские и женские генеративные почки образуются за счет вегетативных, поэтому при обильном цветении появляется меньше ростовых побегов и листьев. На плодоношение расходуется не только крахмал и азотистые соединения, запасенные в древесине, но и углеводы, и другие пластические вещества, образующиеся в процессе ассимиляции. Поэтому мелко семенные породы начинают плодоносить раньше, плодоносят почти каждый год и дают много семян.
Хотя крупные деревья дают больше семян, но самые высокие из них могут плодоносить слабее, благодаря чему больше пластических веществ расходуется на прирост древесины. Это наблюдение имеет особое значение при выборе обсеменителей и формировании семенных плантаций. Но не следует отдавать предпочтение и обильно плодоносящим особям, так как это свойство может сохраниться в последующих поколениях, а лесовод заинтересован в создании лесов имеющих повышенный прирост древесины.
Систематическое семеношение древостоев осины, березы, граба начинается с 15-30 лет, ясеня, липы, ильмовых, лиственниц с 25-40 лет, сосны с 30-50 лет, ели с 40-70 лет, дуба с 40-80 лет, кедра с 50-80 лет. Позднее семеношение резко увеличивается.
Годы обильного урожая семян (семенные годы) в различных климатических условиях и у различных пород повторяются через год у граба; 1-2 года у клена остролистного, через 2-3 года, у ольхи; ясеня через 2-4 года, у сосны, ели, лиственницы через 3-7 лет; бука-5-10 лет. Строгой периодичности нет.
Мелкие семена распространяются дальше. (Ветром у осины на 1-2 км, березы на 0,5-1,0 км). Поэтому древесные породы с мелкими семенами способны возобновляться лучше.
Экологические особенности. Под пологом леса преимущество в возобновлении имеют теневыносливые древесные породы (ель, пихта, кедр, бук и другие), а на не покрытых лесом землях - виды устойчивые к заморозкам (береза, осина, липа, сосна и другие).
Температура воздуха и влажность. На начальные этапы возобновления (цветение и семеношение) влияет главным образом погода. Низкие температуры в начале вегетационного периода приводят к разновременному образованию и развитию мужских и женских цветков, к снижению жизнеспособности пыльцы.
Последующее в вегетационный период охлаждение воздуха до положительной температуры 2-30С может повлечь за собой неурожай семян ели.
Замечено, что во время вегетационных периодов с высоким дефицитом влажности воздуха в июне-июле, и особенно в засушливые годы закладывается большое число генеративных почек. При благоприятных условиях на следующий год наблюдается обильное семеношение хвойных пород.
В сухой почве (подстилке) семена не прорастают. При влажности супеси ниже 10% и подстилки ниже 20% всходы появляются редко.
Оптимальная температура для прорастания семян 18-200С, минимальная 50С. При отсутствии снежного покрова вымерзают крупные семена (желуди и другие).
Освещенность. Малая освещенность под пологом леса ограничивает возобновление даже теневыносливых пород. Буковый самосев появляется при сомкнутости древесного полога 0,9 и ниже. В этих условиях высоты 2м достигает не более 1тысячи экземпляров на 1 га. При сомкнутости 0,8 подрост растет выше (до 3-4 м). Оптимальной считается величина сомкнутости 0,6.
Наибольшее количество елового подроста встречается при полноте 0,6- 0,7. У 3-5 летнего дуба при освещенности 2-10% обмирает или верхушечная почка, или верхние побеги, или все растение.
Лесная подстилка. Лесная подстилка и лесной опад имеют большое значение для лесовозобновления. На прорастание семян и формирование всходов оказывает влияние состав опада и подстилки, ее плотность и толщина, степень разложения, содержание влаги. В зоне подстилки укореняются всходы, закладываются начальные этапы формирования корневых систем будущих деревьев. Мощная и плотная подстилка препятствует прорастанию семян, обеспечению всходов влагой и пищей, проникновению корней в минеральную часть почвы. Более благоприятно воздействует листва, способная к скручиванию, образующая рыхлую подстилку (береза, рябина, ольха, молодая осина и другие). Неблагоприятен для возобновления еловый опад.
Захламленность. Обычная захламленность в лесу благоприятствует лесовозобновлению. На вырубках и гарях захламленность ослабляет физическое испарение и силу заморозков. Как показали исследования
Мелехова, в тайге при захламленности поверхности гари 30-50% ель возобновляется наилучшим образом. Для возобновления сосны также рекомендуется оставлять порубочные остатки.
Влияние растений. Материнские и деревья других видов отрицательно влияют на появление и рост подроста тем, что задерживают свет, иссушают почву. Живой напочвенный покров во многом определяет среду для прорастания семян и развития всходов.
Влияние живого напочвенного покрова зависит в первую очередь от густоты растений. Большую отрицательную роль в возобновлении леса играют задернители, особенно злаки. Образование дернины физически препятствует прорастанию семян.
Конкурирующее влияние напочвенного покрова проявляется и в борьбе за питательные вещества почвы и за свет.
Живой напочвенный покров оказывает большое влияние на возобновление леса через изменение микроклимата.
Воздействие фауны. Фитофаги – отрицательно влияют на лесовозобновления.
Отдельные представители фауны способствуют распространению главных пород. Сойки, дятлы, белки, мыши заносят семена в другие древостои.
Влияние болезней. Болезни семян и плодов, сеянцев и молодых особей проходят на лесной фитопатологии.
Воздействие огня. Низовой пожар вызывает гибель всходов и подроста под пологом леса. Но, уничтожая живой и мертвый напочвенный покров, огонь создает лучшие условия для появления новых поколений леса.
На вырубках сплошной пал губит подрост, а при его повторении возобновление леса замедляется. Но на свежих почвах, вызывая минерализацию почвы и предотвращая ее зарастание травянистокустарниковой растительностью, способствует лесовозобновлению. Таким образом, при анализе и прогнозе лесовозобновления надо принимать во внимание все факторы.
Подрост принято подразделять в зависимости от его высоты на 3 группы: мелкий (высотой до 0,5 м), средний (высотой 0,51-1,5 м) и крупный (выше 1,5 м).
Предварительный подрост по его устойчивости к изменяющимся условиям среды после сплошной рубки классифицируются на 3 категории качества и состояния: надежный (синонимы – благонадежный, жизнеспособный, световой), сомнительный (переходный) и ненадежный (неблагонадежный, нежизнеспособный, теневой).
Выживаемость подроста зависит от особенностей его размещения по площади. Поэтому различают размещение подроста одиночное, групповое и куртинное. При групповом размещении особи подроста имеют общий полог, а размер группы не превышает 10 м2. Куртина имеет большую площадь общего полога в одном месте.
Одиночный подрост, группы и куртины его могут распределяться равномерно (при встречаемости свыше 65%), не вполне равномерно (40-65%) и неравномерно (при встречаемости меньше 40%).
По густоте предварительный подрост следует разделять на очень густой (больше 10 тыс./га) густой (5,1-10 тыс.), средней густоты (3-5 тыс.) т редкий (10,0-2,9 тыс.). При густоте подроста менее 1 тысячи экземпляров на 1 га, возможной сохранности его без повреждений при рубке леса в 50-60% и послерубочного опада в 10-40% оставшееся число особей не имеет хозяйственного значения.
На вырубках подрост характеризуют по степени жизнеспособности. При этом выделяют следующие категории качества хвойного подроста (по методике Мелехова И.С.):
1. благоприятный в физиологическом отношении, но безукоризненный в техническом;
2. благоприятный в физиологическом отношении, но технически дефективный;
3. сомнительный;
4. нежизнеспособный;
5. сухой.
Так как появление самосева зависит от интенсивности семеношения, то оно оценивается глазомерно. Широко применяется глазомерная оценка семеношения В.Г. Каппера.
Наиболее распространенными методами учета подроста являются глазомерно-таксационный и перечислительный (метод учетных площадок и лент).
Глазомерно-таксационный метод применяется во время лесоустроительных работ при таксации леса. При этом записывают состав подроста по числу жизнеспособных особей, густоту его в тыс. экз. на 1 га, пределы возраста и высоту большинства экземпляров, характер размещения подроста и встречаемость главной породы. Точность определения при глазомерно-таксационном методе ± 30-40 %.
Метод учетных площадок чаще всего применяется под пологом древостоя и в научно-исследовательских целях.
В зависимости от густоты и высоты подроста размер учетной площадки может изменяться от 1 до 25 м2.
При отводе лесосек выделяют места, где подрост отсутствует, то есть его густота менее 1,0 тыс. экз. на 1 га.
На остальной площади закладывают от 30 до 100 площадок. На них проводят сплошной перечет подроста по породам, группам высот и категориям качества. Затем общую густоту всего живого подроста переводят на 1 га и эту величину записывают в лесорубочный билет как требование к лесозаготовителю сохранить в указанных местах не менее 50-70 % экземпляров подроста.
Для оценки возобновления по густоте подроста предложено много различных шкал.
В настоящее время для оценки естественного возобновления действует шкала бывшего Гослесхоза. При глазомерном определении густоты подроста в количестве 8 тыс. шт/га под пологом леса его оценивают по встречаемости.
Смена состава древостоев, называемая сменой пород или сукцессией (от латинского succеssic – приемственность), представляет собой изменение лесного биогеоценоза в связи с переходом от преобладания одной породы (эдификатора) к преобладанию другой. В нашей стране особую остроту приобрела проблема смены хвойных лесов лиственными (осиной, березой) в результате применения сплошных рубок леса.
В.Н. Сукачев развивая классификацию Г.Н. Высоцкого к дигрессивным сменам отнес сукцессии, вызванные рубками, вытаптыванием и другими воздействиями человека, даже посадкой или посевом древесных пород. Такие антропогенные смены объединяются вместе с другими сменами, произошедшими в результате внешних воздействий в экзогенную сукцессию. Смены, идущие в направлении восстановления коренных сообществ, называются демутационными. В них решающее значение имеет медленное изменение биогеоценоза под влиянием самой растительности, те есть эндогенная сукцессия.
Смена пород происходит под влиянием следующих факторов:
1. длительного изменения климата;
2. воздействия засухи, ветра, сильных морозов, вызывающих гибель древостоев;
3. изменения почвы в результате эрозии, поднятия или опускания уровня грунтовых вод, уплотнения почвы в рекреационных лесах и т. д.
4. влияния лесной фауны;
5. воздействия огня;
6. лесоводственных свойств древесных пород;
7. загрязнения атмосферы;
8. рубок леса;
9. другой деятельности человека.
Существуют вековые сукцессии, в результате которых произошла смена пород в послеледниковый период.
Вторым видом смен в лесоводстве является длительные сукцессии, когда в течение 150 лет и более доминирует одна порода, которая сменяется в результате действия внутренних (эндогенных) причин. Так, например сосна может сменяться елью, которая может развиваться под пологом.
Третий вид смен – кратковременные сукцессии, протекающие за менее продолжительный срок, не превышающий жизнь одного поколения леса.
Эрозия почв в горах Кавказа, связанная с пастьбой скота, приводила к смене дуба на граб, а затем граба на грабинник (граб восточный), хорошо переносящий сухость почвы. Поднятие уровня грунтовых вод приводит в понижениях к смене ели на сосну, березу, ольху черную, а на повышениях, наоборот, усиливается смена сосны елью.
Быструю смену пород вызывает лесная фауна. Повышение плотности лосиного стада за пределы 2 головы на 1 тыс. га. приводит к смене сосноволиственных молодняков на березово-осиновые, затем на березовосероольховые. Засухи способствуют быстрой смене ели на дуб.
Широко известна роль огня в смене состава пород. Лесные пожары содействуют возобновлению сосны, лиственницы, березы, а иногда осины, ели, дуба.
Огромную роль на смену пород оказывают рубки леса, лесные пожары и лесоводственные свойства древесных пород.
На территории европейской тайги, где совпадают ареалы сосны и ели, после сплошных рубок и лесных пожаров обычно образуются сосняки трех типов развития.
На верхних торфяниках или на сухих и бедных почвах образуются сосняки с еловым подлеском. На относительно бедных почвах формируются со временем смешанные сосновые насаждения. На богатых почвах происходит смена сосны на ель или другие породы.
Эндогенная (внутренняя) сукцессия (смена) сосны на ель протекает медленно и представляет собой длительный тип смены. Начинается она с того, что под сосняками с примесью березы появляется подрост ели от занесенных семян. Благодаря теневыносливости ель достигает высоты второго яруса, а при отпаде крупных берез – первого яруса. Эти ели начинают плодоносить и возникает новое поколение.
При дальнейшем интенсивном распаде первого яруса освещенность под пологом леса возрастает, и появляется подрост сосны. На первых этапах развития разновозрастного ельника из этого подроста может сохраниться некоторая примесь сосны. Господство ели наступает в третьем столетии.
Смена сосны на ель способствуют выборочные рубки.
Обратная смена ели сосной возможна после ветровала ельника или гибели его от пожара. Ель как тонкокорая порода гибнет от огня в первую очередь. Толстая кора сосны защищает ее от низовых пожаров. Но если сосна повреждена огнем смертельно, то перед гибелью сильно плодоносит. В огне сгорает подстилка, обнажается минеральный слой почвы. В этих условиях семена имеют высокую групповую всхожесть. Всходы сосны меньше страдают от заморозков, недостатка влаги, чем ели. Возникает сосновый молодняк.
Но иногда в результате пожара формируются одновозрастные ельники.
Смена ели березой и осиной
Смена ели березой и осиной происходит в результате резких изменений
в условиях среды под влиянием сплошной рубки, пожара и т. п. и тех преимуществ, которыми обладают береза и осина перед елью в этих условиях. В неблагоприятную сторону для ели уменьшается здесь микроклимат. Появляется опасность заморозков, солнцепека, других отрицательных явлений. Для березы и осины эти условия благоприятны. Световые условия, легкость семян способствуют их разносу на далекие расстояния, быстрый рост – все это облегчает захват территории березой и осиной.
Вследствие развития сплошных рубок на больших площадях выросла масштабность смены ели березой и осиной.
Вюжных районах тайги в подзоне смешанных лесов в процессах смены ели большую роль играет осина. К северу от этих районов роль осины
всмене пород резко снижается. Преобладающее положение в составе лиственных древостоев на месте гарей и сплошных рубок занимает береза.
В южных районах тайги осина может заселять влажные или свежие
почвы.
С образованием сомкнутого лиственного молодняка и его развитием изменяется и среда – обстановка становится лесной с присущими особенностями микроклимата и почвенными условиями. Лиственные породы улучшают почвенно-гидрологические условия. Создаются условия, благоприятные для развития ели. Постепенно ель переходит в первый ярус и вытесняет березу. Это связано и с меньшей продолжительностью жизни березы по сравнению с елью и совершается обычно одним поколением ели, но не всегда.
Скорость восстановления и образования спелого елового леса зависит от размеров площади, наличия и размещения на ней источников обсеменения ели, времени плодоношения, напочвенных изменений, наличия и характера подроста, сохранившегося после рубки и т. д. Длительное преобладание лиственных пород исключает возможность естественной смены их елью на протяжении одного поколения.
Площадь дубрав уменьшается во многих регионах. Смена дуба его спутниками – елью, осиной и березой рассмотрена Г.Ф. Морозовым. Он проследил смену дуба сосной в при степных лесных массивах и указал на возможность обратной смены сосны дубом. Он считал, что сосна заселяет степные почвы, затем под пологом чистого сосняка проявляется дуб с другими породами, формируются многоярусные высокопродуктивные древостои, после сплошной рубки которых происходит смена сосны на порослевой дуб. На определенном этапе развития порослевой дубравы с разреженным пологом в ней появляется самосев сосны, который при определенных условиях перерабатывает порослевой дуб. Лесной пожар ускоряет смену дуба сосной.
В результате сплошных рубок происходит смена дуба малоценными породами. Это происходит во многих регионах, особенно на Северном Кавказе. В результате рубок наблюдается смена дуба и бука на граб и порослевой дуб.
В дубравах северной лесостепи распространена смена дуба осиной. После сплошной рубки даже чистого дубняка наряду с порослевым дубом и его спутником появляется семенная осина. Благоприятными условиями для развития самосева осины являются не только свободные минерализованные места, но и покров из осоки волосистой, кипрея, кусты клена полевого и лещины. С 4-летнего возраста семенная осина дает корневые отпрыски, которые в дальнейшем не отличаются от материнского самосева.
Для восстановления дуба предлагается сплошная рубка при создании культур дуба на вырубках с проведением систематических рубок ухода.
На юге ареала ели европейской многолетние засухи вызвали усыхание ее, и увеличение в составе доли дуба. Однако, если доля дуба превышает 6 единиц, то поселение дубового подроста затрудняется опадом.
Другая картина наблюдается на севере ареала дуба. Так вековые изменения климата приводят к необратимой смене дуба елью. Усиливается смена дуба под влиянием сплошных рубок.
Елово-дубовые леса в результате сплошных рубок сменяются не только осиной и березой, но и ольхой черной. Так как ольха черная не является теневыносливой породой, то предотвращающей эту смену мерой является переход от сплошных рубок к добровольно-выборочным с систематическим уходом за самосевом дуба и ясеня. При отсутствии подроста главных пород на вырубках потребуется создание культур дуба и ели по нетрадиционным технологиям.
Биологическая и экологическая оценка смены древесных пород
Основным фактором смены пород являются сплошные рубки, способствующие распространению осины и березы. В европейской части России смена хвойных пород лиственными произошла не менее чем на 50% площади их рубки, сосняков – на 40%. На громадных площадях прошлых вырубок созревают урожаи менее ценной древесины.
Всегда ли такая смена является нежелательной?
Промышленное значение березы постоянно растет. Древесина березы используется в фанерном производстве, финны изготавливают красивую мебель и создают культуры березы повислой. Березняки имеют высокие рекреационные свойства. Кроме того, береза положительно влияет на почву, ускоряя круговорот веществ. Березовый опад содержит больше азота, кальция, магния, фосфора и быстро минерализуется, увеличивая богатство почвы. В зоне смешанных лесов 30-летнее произрастание березы после сплошной рубки ельника привело к увеличению гумуса на 0,5-1,0%.
Подобное влияние на почву оказывает и осина в молодом возрасте. Она, несмотря на фаутность стволов из-за развития гнили, также иногда полезна. Осина на месте вырубки кисличников, которые могут превратиться в пустыри, если не зарастут в первые 2-3 года характеризуется высокой производительностью. В этих условиях она меньше поражается гнилью и, срубленная в 30-40 лет дает некоторый выход деловой древесины для изготовления бумаги, спичек, паркета, дачных домиков и др. Она является кормом для лося и других животных.
Чистые культуры ели и сосны могут разрушаться от болезней и вредителей. Для ели примесь березы и осины необходима не только для защиты ее от заморозков: при организации сбыта древесины мягколиственных пород эффективнее в экономическом и биологическом плане в первые десятилетия выращивать ель под ярусом березы, осины то есть допускать кратковременную смену пород. Такая смена выгодна потому, что сформировавшийся ельник из осветленного второго яруса будет более продуктивен на почве, обогащенной мягколиственными породами.
Хотя в отдельных случаях смена хвойных пород на мягколиственные допускается, в целом она нежелательна.
Расширение использования древесины мягколиственных пород – вынужденная мера, следствие нежелательной смены лесных биогеоценозов.
Рекомендуются следующие меры, сдерживающие нежелательные смены пород и позволяющие восстановить наиболее продуктивные леса:
1. Борьба с лесными пожарами и выращивание в случае их возникновения на гарях сосновых (при близком залегании мерзлоты – лиственных) насаждений.
2. Правильный выбор способа главной рубки, позволяющего использовать потенциальные возможности самовозобновления хозяйственно ценных пород.
3. Проведение мер содействия естественному возобновлению главных пород, и в первую очередь сохранение подроста при лесозаготовках.
4. Создание подпологовых и других лесных культур.
5. Своевременное проведение рубок ухода или химического ухода в молодняках.
Подробнее эти мероприятия рассматриваются в практическом лесоводстве.
1. Расскажите о методах возобновления леса.
2. Охарактеризуйте виды размножения древесных и кустарниковых пород.
3. Расскажите о способах классификации подроста.
4. Укажите факторы, влияющие на возобновление леса под пологом древостоях, на вырубках и гарях.
5. Укажите основные причины и виды смены состава древостоев.
6. Опишите процесс смены ели березой и осиной.
7. При каких условиях, и в каких типах леса происходит смена сосны елью и ели сосной?
8. Дайте биологическую и экономическую оценку смены хвойных пород березой и осиной.
Морозов Г.Ф. Учение о лесе - М.-Л., 1930.
Мелехов И.С. Лесоведение и лесоводство. - М.: МГУЛ, 1972.
Колесников Б.П. Лесоведение и лесоводство, т.1. - М., 1975.
Мелехов И.С. Лесоводство. - М.: МГУЛ, 2003. - 320 с.
Мелехов И.С. Лесоведение. - М.: МГУЛ, 2004. - 398 с.
Маркова И.С. Лесоведение - Курс лекций, М., 2005.
Сенов С.Н., Грязькин А.В. Лесоведение - СПб, 2006.
Лесоустройство - Конспект лекций по В.Е.Ермакову, Минск, 2010
Захаров А.Ю., Мишин Н.С. Лесоводство - Методические рекомендации к проведению учебной практике по лесоводству, Архангельск, 2015.
Погребняк П.С. Общее лесоводство. М.: Колос, 1968. - 440 с.
Arkhipov V.A., Khaidarov K.A., Moukanov B.M., Goldammer J.G. Overview on Forest Fires in Kazakhstan. - International Forest Fire News No. 22 - April 2000, p.40-48.
Khaidarov K.A., Arkhipov V.A. Forest Fire Situation in Kazakhstan. - International Forest Fire News No. 24 - April 2001, p.60-66.
Флейзор Л. и др. Состояние лесов мира - Рим, 2011.
Астанинская резолюция по восстановдению лесов - Астана, 2018.
1. Мелехов И.С. Лесоведение. М.: МГУЛ, 2004. - 398 с.
2. Бельгард А.Л. Степное лесоведение. - М.: «Лесная промышленность», 1971. - 336 с.
3. Погребняк П.С. Общее лесоводство. М.: Колос, 1968. - 440 с.
4. Тихонов А.С., Набатов Н.М. Лесоведение: учебник для высших учебных заведений. – М.: Экология, 1995.
5. ОСТ 56-108-98 Лесоводство. Термины и определения./ ВНИИЦлесресурс. – М, 1994. - 56 с.